Flowering time of wheat varieties in New South Wales
Author: Martin P.J., Eagles H.A., and Cane K. | Date: 04 Mar 2014
Varieties displaying this symbol beside them are protected under the Plant Breeders Rights Act 1994.
Authors
Martin P.J., Eagles H.A., and Cane K.
Take home message
It is critical to match variety and sowing date so that flowering occurs early enough to allow a long grain filling period before the high evaporative demands and soil water deficit of early summer. The flowering period must also be late enough to avoid damage by frosts in early spring.
Because the timing of the autumn break can range from April until June, farmers in NSW have the choice of a range varieties with differing maturities. Understanding how each variety responds to the environment will help target varieties to their best sowing time.
Maturity
The relative maturity of varieties is controlled by responses to photoperiod, vernalisation and earliness per se.
Photoperiod response is a plant’s response to day length (the number of hours of daylight). Wheat responds to long days. This means that lengthening days induces a wheat plant to change from vegetative to reproductive development.
Vernalisation is a plant response to cold temperature. Vernalisation promotes the transition from vegetative to reproductive development. When vernalisation has been satisfied, apical meristems change from producing leaf primordia to producing floral primordia. In wheat, vernalisation accumulates most rapidly between about 3°C and 10°C.
Earliness per se, which is also called propensity to flower, has been defined as the period from floral initiation (double ridge, GS30) until flowering independently of photoperiod and vernalisation.
Accumulated growing day degrees has an effect on the overall length of the growing period. This effect is the same for all varieties and therefore does not change the relative maturity of varieties.
Genetic controls
The genetic control of maturity is complex. All 21 chromosomes of wheat reported to influence maturity in some way. The development of diagnostic markers for the photoperiod gene Ppd-D1, and the vernalisation genes Vrn-A1, Vrn-B1 and Vrn-D1has allowed the characterisation of current commercial varieties ( Eagles, Cane, & Vallance, 2009) and Table 1.
The photoperiod genes Ppd-B1 and Ppd-D1, and the vernalisation genes Vrn-A1, Vrn-B1 and Vrn-D1 have been shown to account for approximately 53% of the genetic variance for days to heading in southern Australia (Cane et al., 2013).
The vernalisation and photoperiod genes interact with each other in an epistatic manner, meaning that the effect on vernalisation requirement is much greater when genes are present in combination (Table 1).
|
Combination of allelesA |
Days to heading |
Typical variety |
|---|---|---|
|
aaa |
116.4 |
Catalina |
|
vaa |
116.8 |
Drysdale |
|
difference |
-0.4 |
|
|
sed |
0.7 |
|
|
ava |
116.0 |
H46 |
|
vva |
118.3 |
EGA Gregory |
|
difference |
-2.3* |
|
|
sed |
0.7 |
|
|
avv |
119.4 |
Janz |
|
vvv |
123.2 |
EGA Wedgetail |
|
difference |
-3.8* |
|
|
sed |
0.6 |
|
From Table 1, the difference in days to heading between genotypes differing only at the Vrn-A1 locus is greatest when winter alleles are present at Vrn-B1 and Vrn-D1 (avv versus vvv), intermediate when a spring allele is present at Vrn-D1 (ava versus vva) and negligible when spring alleles are present at both Vrn-B1 and Vrn-D1 (aaa versus vaa). Typical varieties are presented for illustrative purposes, but these comparisons were made using data across southern Australia (including southern NSW, Victoria and South Australia) for many varieties and breeding lines.
There are a large number of other genes which contribute to maturity. Some of these are known to occur in Australian varieties, for example Vrn-D4, the winter allele is present in the winter varieties currently grown in New South Wales, while the spring allele occurs in the facultative variety Oxley. Some are currently not possible to characterize given the current availability of diagnostic markers.
It is difficult to determine which genes are present without diagnostic markers for the various alleles concerned. However, an increasing number of diagnostic markers are being developed by research programs in Australia and around the world. As these markers become available and the current suite of available varieties is characterized our understanding of the drivers of maturity will gradually be improved.
How are wheats classified?
Based on the response of varieties to vernalisation and photoperiod they can be categorised as either a winter, facultative spring or spring wheat (Table 1). Within each category there is a range in flowering dates between varieties.
Varieties with the vernalisation genes (Vrn-A1, Vrn-B1 , Vrn-D1 and Vrn-D4) are classified as winter wheats. Examples are EGA Wedgetail, Rosella, Whistler and Wylah.
Varieties with combinations of the photoperiod and only one or two Vrn vernalisation genes fit into the facultative and spring categories.
Figure 1 shows how some of the current varieties are classified.
Figure 1. Estimated response of wheat varieties to vernalisation and changes in photoperiod.
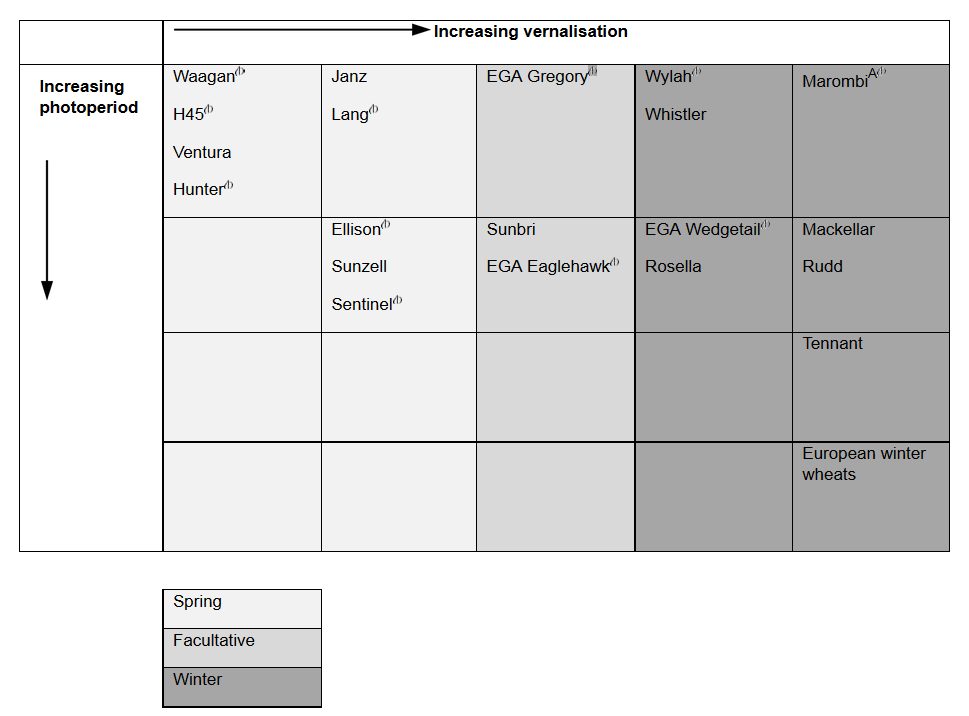
Figure 1 text description: The Spring wheat varieties showed a fairly even estimation of vernalisation to photoperiod and at quite low levels for each. The wheat varieties Waagan, H45, Ventura and Hunter all showed the lowest possible vernalisation and photoperiod. Facultative wheat varieties had a slightly higher vernalisation than photoperiod, with vernalisation being around the mid level and photoperiod staying around the lower levels. Winter wheat varieties showed a mostly high vernalisation to low photoperiod. However, European winter wheats had both the highest vernalisation and photoperiod possible and Tennant had a very high vernalisation to high photoperiod.
Note A- Marombi has a different and stronger Vrn-A1 allele than other Australian winter wheats, meaning that it takes longer to vernalise in winter. However, once vernalised it responds rapidly to increasing temperature and photoperiod, and does not head later than MacKellar and Rudd.
|
Cultivar |
Genes | Maturity group in NSW |
||||
|---|---|---|---|---|---|---|
|
Ppd-B1 |
Ppd-D1 |
Vrn-A1 |
Vrn-B1 |
Vrn-D1 |
||
|
Axe |
a |
a |
a |
a |
v |
S |
|
Barham |
a |
d |
a |
a |
v |
S |
|
Beaufort |
b |
a |
b |
v |
v |
F |
|
Bolac |
b |
a |
a |
v |
v |
F |
|
Braewood |
a |
d |
a |
v |
v |
F |
|
Carinya |
c |
a |
a |
v |
v |
S |
|
Catalina |
b |
a |
a |
a |
a |
S |
|
Chara |
b |
a |
a |
v |
v |
S |
|
Corack |
b |
a |
v |
a |
a |
S |
|
Correll |
a |
d |
a |
a |
v |
S |
|
Crusader |
d |
a |
v |
a |
a |
S |
|
Cunningham |
c |
a |
a |
v |
v |
F |
|
Currawong |
b |
a |
v |
v |
v |
W |
|
Dart |
b |
a |
a |
a |
v |
S |
|
Derrimut |
d |
a |
a |
v |
v |
S |
|
Diamondbird |
b |
a |
v |
a |
v |
S |
|
Drysdale |
b |
a |
v |
a |
a |
S |
|
EGA_Bounty |
b |
a |
v |
v |
a |
S |
|
EGA_Eaglehawk |
b |
b |
b |
v |
a |
F |
|
EGA_Gregory |
b |
a |
v |
v |
a |
F |
|
EGA_Stampede |
a |
a |
v |
a |
a |
S |
|
EGA_Wedgetail |
b |
a |
v |
v |
v |
W |
|
EGA_Wills |
a |
a |
v |
v |
a |
S |
|
Elmore |
#N/A |
#N/A |
#N/A |
#N/A |
#N/A |
S |
|
Ellison |
a |
b |
v |
a |
a |
F |
|
Emu_Rock |
b |
a |
a |
a |
v |
S |
|
Espada |
a |
d |
a |
a |
v |
S |
|
Estoc |
b |
d |
a |
a |
v |
S |
|
Forrest |
b |
c |
a |
a |
v |
F |
|
Gauntlet |
a |
a |
a |
v |
v |
S |
|
Gazelle |
a |
d |
|
a |
v |
S |
|
Giles |
d |
a |
a |
v |
v |
S |
|
Gladius |
a |
d |
a |
a |
v |
S |
|
Guardian |
d |
a |
a |
v |
v |
S |
|
H45 |
d |
a |
a |
v |
a |
S |
|
H46 |
d |
a |
a |
v |
a |
S |
|
Hartog |
b |
a |
v |
a |
a |
S |
|
Impala |
b |
a |
a |
v |
v |
S |
|
Janz |
c |
a |
a |
v |
v |
S |
|
Kennedy |
b |
a |
v |
v |
a |
S |
|
Lancer |
a |
a |
a |
v |
v |
S |
|
Lang |
a |
a |
a |
v |
v |
S |
|
Leichhardt |
b |
a |
v |
a |
a |
S |
|
Lincoln |
b |
a |
a |
a |
v |
S |
|
Livingston |
b |
a |
a |
a |
v |
S |
|
Marombi |
a |
b |
w |
v |
v |
W |
|
Mackellar |
|
a |
v |
v |
v |
W |
|
Merinda |
b |
a |
a |
v |
v |
S |
|
Merlin |
b |
a |
v |
a |
a |
S |
|
Naparoo |
b |
a |
v |
v |
v |
F |
|
Orion |
a |
d |
a |
a |
v |
S |
|
Oxley |
d |
a |
v |
v |
v |
S |
|
Phantom |
a |
c |
v |
a |
a |
S |
|
Rosella |
a |
d |
v |
v |
v |
W |
|
Rudd |
|
a |
v |
v |
v |
W |
|
Scout |
b |
a |
v |
a |
a |
S |
|
Sentinel |
a |
c |
v |
a |
a |
S |
|
Shield |
a |
a |
v |
a |
v |
S |
|
Spitfire |
a |
a |
v |
a |
a |
S |
|
Strzelecki |
b |
a |
v |
v |
a |
S |
|
Sunbri |
a |
c |
a |
v |
v |
F |
|
Sunbrook |
b |
b |
b |
v |
a |
F |
|
Sunco |
a |
a |
a |
v |
v |
S |
|
Sunguard |
a |
a |
a |
v |
v |
S |
|
Suntop |
d |
a |
v |
a_v |
a |
S |
|
Sunvale |
a |
a |
a |
v |
v |
F |
|
Sunvex |
a |
a |
a |
v |
v |
S |
|
Sunzell |
b |
b |
v |
a_v |
a |
S |
|
Trojan |
a |
c |
v |
a |
a |
S |
|
Ventura |
b |
a |
v |
a |
a |
S |
|
Waagan |
c |
a |
v |
a |
a_v |
S |
|
Wallup |
b |
a |
a |
a |
v |
S |
|
Whistler |
d |
a |
v |
v |
v |
W |
|
Wylah |
a |
a |
v |
v |
v |
W |
|
Yenda |
a |
d |
b |
a |
v |
S |
|
Yitpi |
b |
d |
a |
a |
v |
S |
Gene characterisation data were sourced from (H.A Eagles et al., 2009), with further characterisation conducted by Karen Cane and Jayne Wilson. See (Cane et al., 2013; Eagles et al., 2010) for more information.
The effect on sowing time
Responses to a range of sowing times of varieties which represent winter or facultative and spring genotypes are shown in Figure 2.
When sown in a period of the year with long days and warm temperatures, when vernalisation is not being satisfied, spring varieties and some facultative varieties flower very early. Winter wheats, however, will not flower until their vernalisation requirement is met.
As sowing is progressively delayed into the cooler period of the year, varieties requiring vernalisation, i.e. winter varieties and the facultative and spring varieties flower much more closely together. For example when sown in 2006 on 10/3/2006 Janz flowered 115 days before EGA Wedgetail, when sown in on 10/5/2006 Janz flowered 6 days before EGA Wedgetail
(Figure 2).
Figure 2. Flowering times of wheat varieties sown at a range of sowing times at Temora in 2006.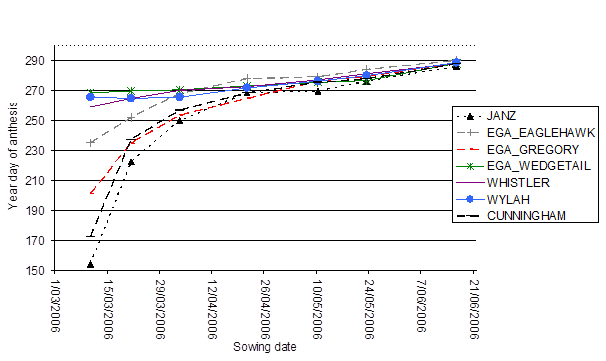
Figure 2 text description: All year days of anthesis increases with a later sowing date. The sowing dates of approximately 10th March till approximately the 5th April have a big impact on the flowering times for Janz, EGA Eaglehawk and EGA Gregory. All three varieties have a very early flowering time early in the year but as the sowing date increases the flowering time makes a large jump to later in the year day of anthesis. After approximately 5th April, these three varieties have a steady increase of year day of anthesis as the sowing date increases. All other varieties flowering date steadily increases later in the year as the sowing date becomes later in the year. All varieties show a similar day of anthesis of approximately 285 when sown on 21st June.
Notes: solid lines are winter varieties, long dashed lines are facultative and dotted lines are spring.
Eagles et al. (2010) suggested that genotypes with only a spring allele at Vrn-D1 respond more to vernalisation than those with only a spring allele at Vrn-A1, but less than winter varieties, and in these time of sowing experiments EGA Gregory![]()
showed an intermediate response to time of sowing compared to Janz and EGA Wedgetail![]() .
.
References
Cane, K., Eagles, H.A., Laurie, D.A., Trevaskis, B., Vallance, N., Eastwood, R.F., . . . Martin, P.J. (2013). Ppd-B1 and Ppd-D1 and their effects in southern Australian wheat. Crop and Pasture Science, 64, 100-114.
Eagles, H. A., Cane, K., Kuchel, H., Hollamby, G. J., Vallance, N., Eastwood, R. F., . . . Martin, P. J. (2010). Photoperiod and vernalization gene effects in southern Australian wheat. Crop and Pasture Science, 61(9), 721-730. doi: doi:10.1071/CP10121
Eagles, H.A, Cane, K., & Vallance, N. (2009). The flow of alleles of important photoperiod and vernalisation genes through Australian wheat. Crop and Pasture Science, 60, 646-657.
Contact details
Peter Martin
Howqua Consulting
Ph: 0418 972 105![]()
Email: p.martin56@bigpond.com
Was this page helpful?
YOUR FEEDBACK