Tolerance to ion toxicities enhances grain yield in acid soils prone to flooding and terminal drought
Author: H Khabaz-Saberi, Z Rengel and S Barker, University of Western Australia | Date: 25 Feb 2015
Key messages
- Ion toxicities induced in transiently waterlogged acid soils reduce root and shoot growth of intolerant wheat genotypes
- Drought accentuates the adverse effect of ion toxicities on root growth>
- Wheat genotypes with enhanced tolerance to ion toxicities produce more grain yield than intolerant genotypes in acid soils prone to transient waterlogging and terminal drought
Aim
Tolerance to toxicity of Al, Mn or Fe resulted in enhanced root and shoot growth during transient waterlogging. This study tested whether such genotypes would also be resilient to drought by growing more extensive and deeper roots and exploiting water resources deeper in the acid soil profile.
Method
The experiments were carried out: (i) at three experimental sites: Merredin Research Station (MERS), Wongan Hills Research Station (WHRS) and Esperance Downs Research Station (EDRS) and (ii) in a phytotron (day–night temperatures set at 20°C-15°C) at the University of Western Australia.
Selection of genotypes
Selection of genotypes contrasting in tolerance to ion toxicity but with similar maturity for the field study was conducted in the phytotron. The uniform seeds of genotypes and reference wheat genotypes of known tolerance: ET8 (Al-tolerant) and ES8 (Al-intolerant), Norquay (Mn-tolerant) and Columbus (Mn-intolerant), Siete Cerros (Fe-tolerant) and BH1146 (Fe-intolerant) were used in the screening. Tolerance to each toxicity was tested separately in all the genotypes.
In screening for Al tolerance, a modified root growth method was employed to determine relative root length. Germinated seeds were grown in the aerated nutrient solutions containing 0 (control) or 440 µM Al as AlCl3. The solutions were maintained at pH 4.2 with daily adjustments as needed. After 7 days, the length of the main seminal root was measured on five seedlings per pot. Aluminium tolerance was expressed as the relative root length (Al treatment/control).
For Mn tolerance assay, seedlings were pre-treated at 2 µM Mn; 7-day-old seedlings were mounted over pots containing aerated nutrient solutions with Mn treatments: 2 µM Mn (control) or 750 µM Mn (high-to-toxic). The solution pH was maintained at 4.8 by measuring daily and adjusting as necessary. The solutions were changed weekly; in between changes, the solutions were adjusted periodically to compensate for a water loss by evapotranspiration. After 14 days of Mn treatment, Mn toxicity symptoms were recorded; in addition, root and shoot dry weight was determined after drying at 60 °C until constant weight. The relative shoot, root and whole-plant dry weight of seedlings grown in nutrient solution with 750 µM Mn was calculated with respect to control (2 µM Mn).
For Fe tolerance assay, aerated nutrient solution containing 36 or 625 µM Fe was used. At the latter concentration, maximum discrimination between Siete Cerros (Fe-tolerant) and BH1146 (Fe-intolerant) was observed. The screening conditions and the measured parameters were as described above for Mn tolerance assay, except the solution pH was maintained at 4.5 throughout the assay.
A completely randomised block design with three replicates was employed in all screenings. Data were subjected to analysis of variance (with LSD 5% for comparison among means).
Grain yield evaluation of genotypes contrasting in ion toxicities
Grain yield evaluation of genotypes contrasting in ion toxicities was followed at three acidic field sites in plots with and without lime. The extremely dry conditions at MERS (104 mm during the growing season, ~55% of the long-term mean) and WHRS (119 mm during the growing season, 51% of the long-term mean) and the nearly long-term average rainfall at EDRS (400 mm in-season rainfall, 109% of the long-term mean) during 2010 furnished excellent natural conditions to evaluate performance of genotypes under combined drought and soil acidity.
The experiments were laid out in a randomised strip-plot design (plus and minus lime strips). Lime at 3 t ha–1 was top dressed at WHRS and MERS in 2008 and at EDRS in 2010. Plots contained seven rows (22-cm row spacing) and were 10-m long at sowing (8-m length was harvested for grain yield). There were three replicates at MERS and WHRS sites and four replicates at EDRS site. The replicate soil samples were taken from three depths (0–10, 10–20 and 20–30 cm) and analysed for physical and chemical properties using standard methods. Grain yield and relative grain yield at moisture-limited vs. non-limited site (i.e. grain yield at either MERS or WHRS/grain yield at EDRS) as well as the soil analyses data were subjected to analysis of variance using GENSTAT Release 9.2 (VSN International, Hemel Hempstead, UK).
Results
Selection of genotypes distinct in tolerance to ion toxicities
The selected Australian Al-, Mn- and Fe-tolerant and intolerant genotypes were quite distinct in tolerance to ion toxicities. They were comparable to the international reference genotypes in the case of the ion in question (Fig. 1).
Soil feature, rainfall distribution and lime application
Regarding subsoil, the trial sites were strongly acidic at WHRS and MERS (3.7–3.9 in 0.01 M CaCl2) and relatively mildly acidic at EDRS (4.4–4.7 in 0.01 MCaCl2). A significant increase in the surface soil pH upon addition of lime occurred at all sites (data not shown). With growing-season rainfall at MERS, WHRS and EDRS being, respectively, 55, 51 and 109% of long-term average, moisture availability at MERS and WHRS was limiting during anthesis and post-anthesis, whereas it was adequate at EDRS throughout the growing season. The low rainfall and strongly acidic subsoil at WHRS and MERS compared with the relatively mild subsoil acidity coupled with adequate and uniformly distributed rainfall at EDRS was the basis for assessing the genotypes under combined drought and acidity at MERS and WHRS vs. their yield potential at EDRS.
Grain yield and relative grain yield of genotypes at MERS and WHRS (moisture-limiting sites) and EDRS (adequate moisture)
The grain yield of the genotypes at EDRS, as the site with adequate moisture, and at two drought-affected sites (MERS and WHRS) in the treatments with or without lime (Table 1) and the relative grain yield at MERS or WHRS as a percentage of the yield at EDRS (Fig. 2) were presented. The average yield at EDRS (3618 kg ha–1) was ~3-fold higher than at WHRS (1231 kg ha–1) and ~4.5-fold higher than at MERS (783 kg ha–1).
Liming was not effective in increasing grain yield of genotypes at EDRS (Table 1). At WHRS, however, liming generally increased grain yield of most genotypes (and the liming x genotype interaction was non-significant) (Table 1). At MERS, liming increased grain yield of Mn-tolerant more than Mn-intolerant genotypes; hence, there was a significant liming x genotype interaction (Table 1).
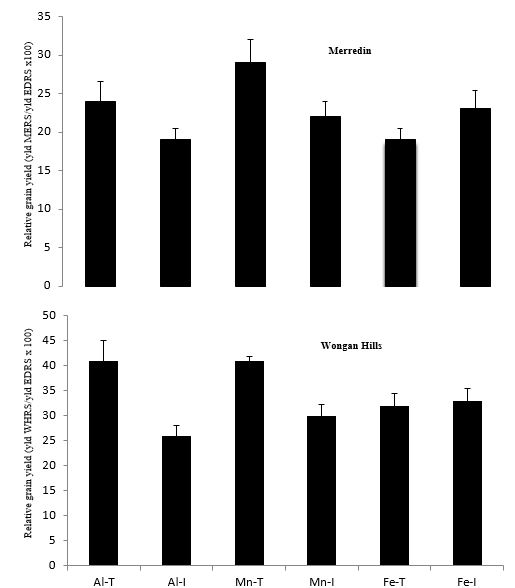
At both WHRS and MERS, Al-tolerant and Mn-tolerant genotypes had higher grain yield than their intolerant counterparts, whereas no significant difference was observed between Fe-tolerant and Fe-intolerant genotypes (Table 1). There were no significant differences in grain yield among genotypes at EDRS. In terms of relative yield (moisture-limited site/moisture-non-limited site), Al-tolerant and Mn-tolerant genotypes outperformed their intolerant counterparts at both WHRS and MERS (Fig. 2). There was no significant difference between Fe-tolerant and Fe-intolerant genotypes (Fig. 2).
Conclusion
The difference in tolerance of the selected wheat genotypes to toxicities of Al (6-fold), Mn (14-fold) or Fe (2.7-fold) (Fig. 1) was in agreement with the previous reports and confirmed the suitability of the selected genotypes contrasting in tolerance to individual ion toxicities for assessing tolerance to multiple ion toxicities that occur in waterlogged acid soils.
This study verified the effectiveness of liming as the chemical approach in combination with the genetic approach based on use of acid- and Al-tolerant genotypes. The positive yield response of genotypes to lime (Table 1) could be explained by partial amelioration of soil acidity in the surface layer where roots were confined early in the season; however, the topsoil, which was wet in the early part of the season, dried up later during anthesis and post anthesis, forcing genotypes to rely on subsoil moisture reserves that could be accessed by deep roots only.
The Al-tolerant and Mn-tolerant genotypes are likely to grow roots into acidic subsoil horizons, thus allowing more water extraction from these deep horizons. The yield potential of genotypes was relatively well expressed at EDRS, where they had higher grain yield in contrast to the dry sites; compared with the EDRS (moisture-non-limited site), relative grain yield at the dry sites was reduced substantially, but more in intolerant genotypes (76%) than tolerant genotypes (68%) on average (Table 1 and Fig. 2). All tested genotypes had a similar yield potential under well-watered conditions and no acidity (no significant yield differences at EDRS, Table 1). Bearing the similar yield potential in mind, the differential yield response and enhanced performance of tolerant genotypes vs. intolerant ones at MERS and WHRS could be attributed to the former having improved uptake of water and nutrients from the surface soil early in the growing season and from acidic subsoil horizons late in the growing season. The adverse effect of drought on grain yield of the genotypes tested was mainly through reduction in number of grains per spike and in seed weight; such reduction was more severe in intolerant than tolerant genotypes (data not shown).
Further work including monitoring root growth of the genotypes (preferably near-isogenic lines differing only in tolerance to ion toxicities) and dynamics of soil moisture through the soil profile is required to ascertain the underlying mechanisms. Given that tolerance to Al or Mn toxicity individually has now been shown to improve grain yield in transiently waterlogged soils, pyramiding tolerances to ion toxicities is a valid approach to widen crop adaptation to acid soils constrained by transient waterlogging followed by drought later in the season.
Acknowledgements
We thank M. Smirk for ICP analyses, and B. Regmi, K. Kariman and P. Damon for invaluable assistance in the experimental work. We are grateful to B. Paynter for sharing ± lime strips (at WHRS and MRS) and S. Dougall, M. Harrod and C. Matthews for running and managing the field trials. The funding for this study was provided by the University of Western Australia.
Table 1. Grain yield of Al-, Mn- and Fe-tolerant and -intolerant hexaploid wheat genotypes grown in ± lime (3 t ha-1 vs. un-limed control) strips at two drought-affected acid soils (Wongan Hills WHRS and Merredin MERS) and the site (Esperance Downs EDRS) with no moisture limitation
| Genotypes | Grain yield (kg/ha) | |||||
|---|---|---|---|---|---|---|
| WHRS |
MERS |
EDRS |
||||
| - lime | + lime | - lime | + lime | - lime | + lime | |
| Al-tolerant | 1258 | 1596 | 833 | 859 | 3705 | 3281 |
| Al-intolerant | 997 | 1018 | 697 | 727 | 3677 | 3968 |
| Mn-tolerant | 1172 | 1634 | 859 | 1166 | 3283 | 3536 |
| Mn-intolerant | 987 | 1164 | 864 | 768 | 3341 | 3941 |
| Fe-tolerant | 960 | 1487 | 667 | 788 | 3747 | 3859 |
| Fe-intolerant | 1144 | 1359 | 848 | 889 | 3536 | 3987 |
WHRS; Lsd 5% lime: 129, genotype: 259, lime x genotype: ns
MERS; Lsd 5% lime: 84, genotype: 177, lime x genotype: 237
EDRS; lime: ns, genotype: ns, lime x genotype: ns
Fig. 1. Response of wheat genotypes contrasting in (A) Al, (B) Mn and (C) Fe tolerance compared with the reference genotypes (ET8: Al-tolerant; ES8, Al-intolerant; Norquay, Mn-tolerant; Columbus, Mn-intolerant; Siete Cerros, Fe-tolerant; BH1146, Fe-intolerant) grown in nutrient solutions with high-to-toxic concentration of Al, Mn or Fe.
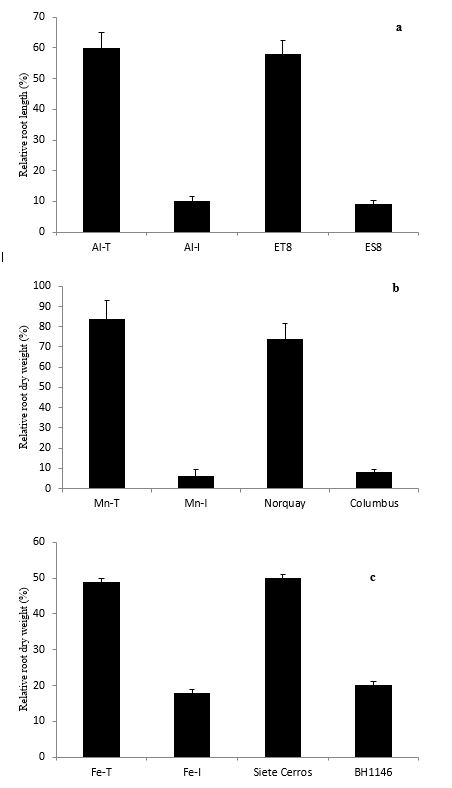
Fig. 2 Relative grain yield of Al-, Mn- and Fe-tolerant (T) and -intolerant (I) genotypes at Merredin (MERS) and Wongan Hills (WHRS) Research Stations (moisture-limited sites) compared with their grain yield at Esperance Downs Research Station (EDRS) (no moisture limitation = 100%).
Was this page helpful?
YOUR FEEDBACK