Weed seed banks – how long do they last?
Author: Dr Catherine Borger, Dr Abul Hashem | Date: 11 Feb 2020
Key messages
- Three to four years of seed-set control are needed to remove a great brome or barley grass seed bank.
- Small seeded species with broad seed dispersal had over 95% of seed emerge in the first year after seed production, while weed species with hard, woody seed coats like Afghan melon, caltrop, doublegee and wireweed had long term seed banks that last at least three years.
- With adequate moisture, all weed species can emerge during the winter-spring growing season; even those species thought of as ‘summer weeds.’ More research is required to control the ‘summer weeds’ like caltrop when they emerge before harvest.
Aims
The key to successful weed management is long-term reduction of the weed-seed bank. However, this requires an understanding of how long the seed bank lasts, which can be influenced by ecotypic differences between weed populations or the environment of the plant producing the seed. For example, Gill and Fleet (2012) noted that barley grass in a cropping system had delayed emergence compared to barley grass on the roadside of the field. The population in the field had evolved delayed emergence to avoid pre-seeding or pre-emergent herbicides. Steadman et al (2004) noted that annual ryegrass grown at warm temperatures produced fewer seeds, and seeds were smaller and less dormant than seeds from plants grown in cool temperatures. Plants in cool, reduced moisture conditions produced seeds that lost dormancy faster than seeds from well-watered plants.
A range of emerging weed species in Western Australia are becoming increasingly common, including Afghan melon (Citrullus amarus Schrad.), barley grass (Hordeum leporinum Link), great brome (Bromus diandrus Roth), button grass (Dactyloctenium radulans (R.Br.) P.Beauv.), caltrop (Tribulus terrestris L.), double gee (Rumex hypogaeus T.M.Schust. & Reveal, syn. Emex australis Steinh.), roly poly (Salsola australis R.Br.), sowthistle (Sonchus oleraceus L.), windmill grass (Chloris truncata R.Br.) and wireweed (Polygonum aviculare L.). Yet we have very little information on seed bank dynamics of these species. The current research aimed to investigate seed dormancy in these ten target weed species, including:
- Peak emergence time and number of years the seed bank can last in controlled conditions and in field conditions (for the winter weed species).
- Influence of maternal environment of the plant (i.e. high or low rainfall environment) on emergence and dormancy.
- Influence of management practices (i.e. plants growing in a crop field or roadside) on emergence and dormancy.
Method
Screen house trial
From Nov 2015 to Jan 2016, seed was harvested from Afghan melon, barley grass, great brome, button grass, caltrop, double gee, roly poly, sowthistle, windmill grass and wireweed populations. Seed from each species was taken from three populations in cropped fields and matching populations on the adjacent roadside, in both high and low rainfall zones of the WA wheatbelt (12 populations per species). In Apr 2016, trays of 30cm by 30cm by 10cm were filled with sand to within 2cm of the top. Seeds (50 seeds of large-seeded species or 100 seeds of small-seeded species) of each population were placed in trays. Trays were placed in an irrigated screen house, in a randomised block design with three replications. All emerged seedlings were counted and removed weekly, from May 2016 to Apr 2017. Note that in the presentation of results, weed emergence is presented by year, where year one is May 2016 to Apr 2017, year two is May 2017 to Apr 2018 and year three is May 2018 to Apr 2019. Each year incorporates a winter and summer growing season to include emergence of winter or summer annual weeds.
An ANOVA was used to analyse the data, with species, location (crop or roadside) and rainfall (high or low rainfall zone) as factors and tray replication as block (VSN International, 2019). Variates included emergence per month as a percent of total emergence in year one (but not year two or three as monthly emergence was too low for valid analysis) and emergence per year as a percent of total emergence. A square root transformation was applied to the emergence data to ensure normal distribution of the residuals. Back-transformed means are presented in the results. The LSD was back transformed as a range, and the maximum LSD is presented in the results.
Field trial
A trial was established at the DPIRD Wongan Hills Research Station (-30.8461, 116.7442) in 2016 and continued until 2019. In 2016, a site with few weeds was selected and non-selective herbicides/autumn tickle were used to clear existing weeds in autumn. Seeds of two populations (crop and adjoining roadside) of great brome, barley grass, windmill grass, doublegee and wireweed were taken from the collections made in ‘Screen house trial’ (ten populations in total). On 26 May 2016, 1000 seeds of each population were manually spread in an area of 1m2. Directly after spreading the weed seeds, wheat cv. Impress Clearfield® Plus was sown, using minimum tillage (knife points and press wheels), at 80kg/ha, 22cm crop row spacing, at a depth of 3-4cm. MacroPro Plus® at 80kg/ha was banded approximately 3cm below the seed. In Jul 2016, 50L/ha Flexi-N® was applied. The trial was established as a completely randomised block design, with four replications (plots of 2m by 5m). After crop emergence and then after each rainfall event during the 2016 season, all weeds were counted and removed. Intervix® at 750mL/ha (inazamox/imazapyr 33/15 g a.i./L, SC, BASF Australia Ltd.) was applied to kill additional weeds in the trial after the target weed seedlings were assessed. Any additional weeds (i.e. species not part of the trial) were hand weeded from the target 1m2 plots where weed seeds were established. The trial was treated with pesticide where necessary to ensure normal crop growth and harvested on 1 Dec 2016.
In 2017, 2018 and 2019, the site was assessed before applying non-selective herbicide, and seedlings of the target weed species were counted and removed. Spray.Seed® at 2.5L/ha (paraquat/diquat 135/115 g a.i./L, SC, Syngenta Australia Pty. Ltd.) was applied directly before seeding on 25 May 2017, 29 May 2018 and 14 Jun 2019, with the seeding system and trial management as for 2016. In Dec 2017, weed seeds were harvested from the same locations that each of the ten populations originally came from, and on 29 May 2018, 1000 seeds were sown in a 1m2 area that was 2m away from the original 1m2 area in each plot. In 2018, when sowing the weed seeds for a second time, all weed seeds were spread directly before seeding the crop. The exception was windmill grass seed, which was spread directly after the crop was sown. In 2018 and 2019, all seedlings were counted and removed from the two separate quadrats per plot.
An ANOVA was used to analyse the data, with weed species, population within weed species and year as factors and number of seedlings per year as the variant (VSN International, 2019). Separate analysis was applied to the seedlings from the seed planted in 2016, and the seed planted in 2018. A square root transformation was applied to both data sets to ensure normal distribution of the residuals. Back-transformed means are presented in the results, again with the maximum LSD value from the back-transformed range.
Results
Screen house trial
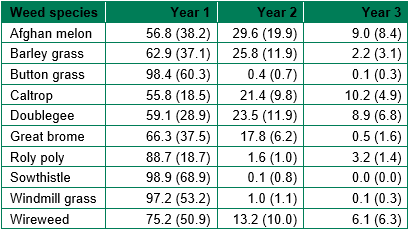
Emergence was affected by weed species, year, and the interaction of species and year (Table 1). All species had highest emergence in year one, and species with small seeds like button grass, roly poly, sowthistle and windmill grass had very low emergence in year two and three. Compared to these species, barley grass and great brome had higher emergence in year two. Hard seeded species like Afghan melon, caltrop, doublegee and wireweed had greater emergence in year two and three. None of the species had all seed germinate over the three years (with germination as a percent of total seed indicated in brackets in Table 1). The trial ended at three years and we did not check if the remaining seeds were dormant or non-viable. There was no impact of population collection site (roadside or cropping field) or rainfall zone (high or low rainfall) on emergence.
Table 1. Average emergence of seedlings as a percent of total emergence over three years (where year 1 to 3 indicates May 2016 to Apr 2017, May 2017 to Apr 2018 and May 2018 to Apr 2019), from seed planted in irrigated conditions in May 2016 (P<0.001, LSD: 12.89). The data in brackets indicate emergence as a percent of total seed planted in each tray.
Great brome, barley grass and doublegee emerged in May-Jul in each year (Figure 1). Button grass, sowthistle and windmill grass had peak emergence in spring (Nov, Sep and Oct) in year one and very low emergence in following years. Wireweed, depending on the year, had peak emergence from Jun to Sep. Afghan melon had peak emergence in Oct (and Aug in year three) and caltrop in Nov. Roly poly emergence was variable between years, with peak emergence in Oct-Nov in year one, Jul in year two and Jun and Sep in year three.
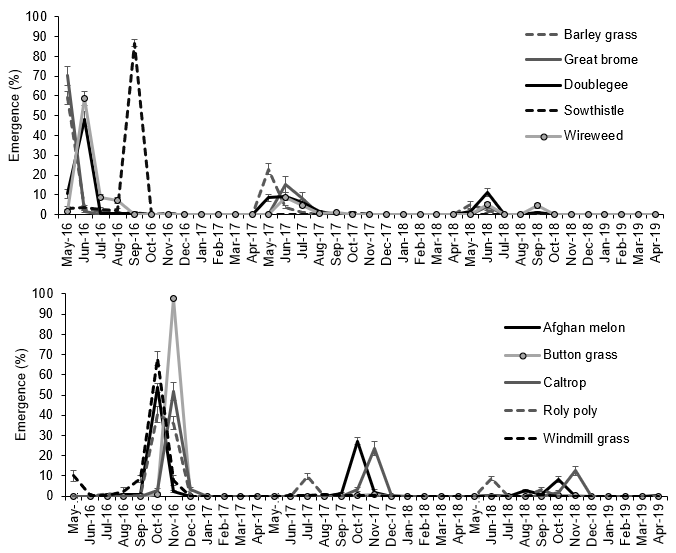
Figure 1. Emergence (as a percent of total emergence) of winter (top) and summer (bottom) weeds in irrigated conditions over three years. The vertical bars indicate the standard error of 32 replications.
Field trial
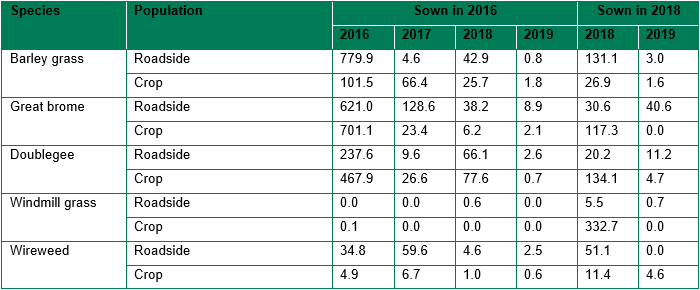
For seed planted in 2016, average emergence of barley grass, great brome and doublegee over all years was much higher than that of windmill grass or wireweed (60.4 barley grass, 101.7 brome grass, 65.4 doublegee, 0.02 windmill grass and 8.9 wireweed, P<0.001, LSD: 11.90). For seed planted in 2018, there was no difference between emergence for each species, when the data is averaged over all years. Windmill grass emergence was much higher when the seed was spread after seeding the crop (2018) rather than before seeding (2016).
For seed planted in both 2016 and 2018, emergence (averaged over all species) was much greater in the first year than subsequent years. For the seed planted in 2016, there were 181.7 seedlings in 2016, 20.3 in 2017, 17.0 in 2018 and 1.3 in 2019 (P<0.001, LSD: 14.37). For the seed planted in 2018, there were 64.3 seedlings in 2018 and 3.2 in 2019 (P<0.001, LSD: 5.74).
There was a significant interaction of weed species, population (roadside or cropped field) and year for both the 2016 and 2018 planted seed, but the effect of population was not consistent between species (Table 2). The windmill grass seed planted in 2018 on the soil surface after sowing the crop had much higher emergence than the windmill grass spread before sowing the crop and incorporated by seeding in 2016. Species generally had much lower emergence in the second year compared to the first year, and the 2016 seed had uniformly low germination in the fourth year. The exception was wireweed, as the seed planted in 2016 had greater emergence of both populations in the second year (2017) compared to the first year (2016).
Table 2. Total average emergence of seedlings (from 1000 seeds) from each weed species population in each year, from seed planted in 2016 (P<0.001, LSD: 93.1) or 2018 (P: 0.004, LSD: 116.10).
Within an individual year, barley grass populations always emerged later than great brome (barley grass peak emergence in Jun/Jul and great brome peak emergence in May/Jun, data presented for seed planted in 2016, Figure 2). For both species, most seedlings had emerged prior to Aug, but some seedlings emerged in Aug, Sep and Oct. Doublegee and wireweed also had staggered emergence throughout the year, although wireweed emergence was low in all years.
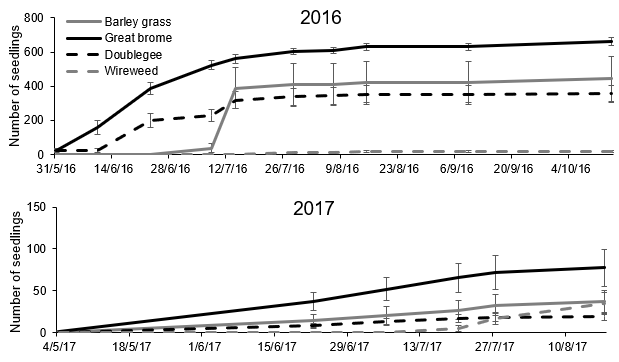
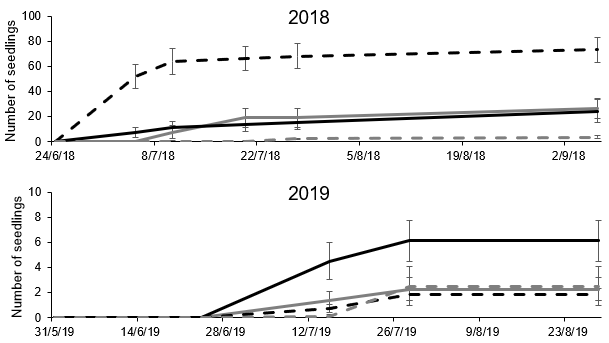
Figure 2. Cumulative emergence of barley grass, great brome, doublegee and wireweed seedlings over the 2016, 2017, 2018 and 2019 cropping seasons, averaged over population, from seed planted in 2016. Vertical lines indicate the standard error of eight replicates.
Conclusion
In irrigated conditions, weed species with small seeds, capable of broad scale wind or water-mediated seed dispersal like button grass, roly poly, windmill grass and sowthistle had high emergence in the first year and very low emergence in subsequent years. By comparison, species with large woody seeds like Afghan melon, caltrop and doublegee had a greater proportion of seed remaining dormant. This is a common ecological trend, as species with broad-scale seed dispersal ensure that seeds reach a range of different environments in the following year, and so do not need dormant seed to protect against dry seasons (Rees, 1993). In the current trial, the seed was not assessed for viability at the end of three years, but it is likely that some of the hard seeded species would have continued to emerge over four to five years, as has already been observed for doublegee (Cheam, 1996).
This research confirms prior research indicating that the seed bank of great brome and barley grass can be driven to very low levels with three to four years of control (Storrie, 2014). However, the number of seedlings emerging in the third or fourth year varied due to population, and in some conditions, more than four years may be required for complete control. The barley grass in the current trials had delayed germination compared to great brome in the field, although emergence of both species was earlier in irrigated conditions. In the field seeds are emerging from varying depths and may be partially protected from precipitation. Delayed emergence will make it more difficult to kill weeds with pre-emergent and non-selective herbicides, but the late emerging weeds will also be less competitive in crop. In South Australia, barley grass in crop fields developed increased seed dormancy (compared to roadside populations) to allow seedlings to escape pre-emergent weed control, but delayed emergence is less common in WA (Gill & Fleet, 2012; Borger & Hashem, 2018). The staggered emergence in field conditions ensure that some plants will escape pre-emergence and early in-crop herbicide application, and late season or harvest weed seed control techniques would be required to control surviving plants.
The species in this trial that we classify as ‘summer weeds’ all emerged in spring, or even winter, given adequate irrigation. ‘Winter-spring’ weeds like sowthistle or wireweed had variable emergence times late in the growing season. Controlling these species late in the season is difficult. There are few herbicide options and it is not practical to spray mature crops before harvest. Roly poly, in particular, had variable emergence between years, which has previously been observed for this species (Borger & Scott, 2009). Further research is required to determine successful control measures for weed species emerging late in the growing season.
This research highlighted how sensitive windmill grass is to burial, with good emergence in controlled conditions or when seed is spread after sowing the crop, and very poor emergence when seed is spread prior to sowing the crop and buried by the seeding operation. This confirms the results of Ngo et al (2017), who noted that germination of windmill grass was severely inhibited by dark conditions and seed did not emerge when buried at any depth. It is likely that windmill grass growing in crops originated from seed blowing into the area (after sowing the crop) from plants along the fence line. Therefore, if these fence lines can be kept clear, windmill grass should not be a major weed in cropping systems.
® Registered Trademark.
Varieties displaying this symbol beside them are protected under the Plant Breeders Rights Act 1994.
Acknowledgments
The research undertaken as part of this project is made possible by the significant contributions of growers through both trial cooperation and GRDC investment, the authors would like to thank them for their continued support. We would also like to thank Nerys Wilkins and Peter Gray for technical assistance, Shari Dougall and Bruce Thorpe at the Wongan Hills Research station for running the trial, Andrew van Burgel for statistical advice and Alex Douglas for reviewing this paper.
Contact details
Dr Catherine Borger, Department of Primary Industries and Regional Development
75 York Rd (PO Box 483) Northam WA 6401
0467 816 082
08 9622 1902
catherine.borger@dpird.wa.gov.au
References
Borger CPD & Hashem A (2018) Emergence of great brome grass and barley grass. In: Proceedings Weed Biosecurity - Protecting our Future. 21st Australasian Weeds Conference. (eds S Johnson, L Weston, H Wu & B Auld). 9-13 September, Manly, Australia. 341-344. The Weed Society of NSW, Sydney, Australia.
Borger CPD & Scott JK (2009) The biology of Australian weeds: 55. Salsola australis R.Br. Plant Protection Quarterly 24, 126-137.
Cheam AH (1996) Doublegee (Emex australis Steinh.) seed banks. Plant Protection Quarterly 11, 141-142.
Gill GS & Fleet B (2012) Seed dormancy and seedling recruitment in smooth barley (Hordeum murinum ssp. glaucum) populations in southern Australia. Weed Science 60, 394-400.
Ngo TD, Boutsalis P, Preston C & Gill G (2017) Plant development and seed biology of windmill grass (Chloris truncata) in southern Australia. Weed Science 65, 395-405.
Rees M (1993) Trade-offs among dispersal strategies in the British flora. Nature 366, 150-152.
Steadman KJ, Ellery AJ, Chapman R, Moore A & Turner NC (2004) Maturation temperature and rainfall influence seed dormancy characteristics of annual ryegrass (Lolium rigidum). Australian Journal of Agricultural Research 55, 1047-1057.
Storrie AE (2014) Integrated weed management in Australian cropping systems. Grains Research and Development Corporation, Australia.
VSN International (2019) GenStat for Windows 19th edition. VSN International, Hemel Hempstead, UK.
GRDC Project Code: UA00156,
Was this page helpful?
YOUR FEEDBACK