Soil amelioration – magnitude of crop productivity improvements on hostile subsoils?
Soil amelioration – magnitude of crop productivity improvements on hostile subsoils?
Take home messages
- Deep placement of organic and inorganic amendments increased grain yield in the order of 20 to 40% for three successive years on a sodic subsoil at Rand.
- Deep placement of organic and inorganic amendments increased root growth and soil water use from the deeper clay layers during the critical reproductive stages of crop development.
- Improvements in grain yield with deep placement of organic and inorganic amendments were associated with a reduction in subsoil pH and exchangeable sodium percentage (ESP%) and an increase in microbial activity.
- Genotypic variability in grain yield response of wheat cultivars grown on sodic subsoils has identified varieties and associated traits for enhanced performance and future breeding.
Background
In the Australian grain belt soil constraints are often expressed as lower water use efficiency (WUE) of grain production and are compared with water limited yield, otherwise known as potential yield. The difference between constrained and potential yield is referred to as the yield gap. Among different soil constraints, sodicity is associated with the largest yield gaps across most of the wheat‐cropping areas of Australia, with an estimated yield loss of $A 1300 million per annum (Orton et al. 2018). Sodic soils exhibit a range of physiochemical properties including the presence of high subsoil exchangeable sodium (Na) concentrations which cause soil dispersion leading to poor subsoil structure, impeded drainage, waterlogging, denitrification and high soil strength. These properties restrict the rooting depths of crop species and subsequent water and nutrients extraction (Incerti and O'Leary 1990; Passioura and Angus 2010) leading to significant yield gaps (Adcock et al. 2007) that affect the profitability of cropping systems (Orton et al. 2018).
In southern NSW winter crops commonly have sufficient water supply either from stored soil water or rainfall during the early growth stages. However, the reproductive phase is often affected by water stress or terminal drought and this is thought to be the major cause of variable grain yield (Farooq et al. 2014). The effect of water stress in the reproductive phase is further impacted by shallow root depth induced by subsoil sodicity. Key to improving crop productivity under such a condition is to improve root growth in and through sodic subsoils to enable use of deep subsoil water late in the growing season. Water use at this late stage has a two- to three-fold greater conversion efficiency into grain yield (Kirkegaard et al. 2007) than seasonal average based conversions efficiencies (i.e. 50 – 60kg/mm versus 20 – 25kg/mm, respectively).
While there are large advantages to be gained by improving the soil environment of sodic subsoils, the various amelioration approaches (deep ripping, subsoil manuring, applying gypsum, improved nutrition and use of ‘primer-crops’) have produced variable results (Adcock et al. 2007; Gill et al. 2008). Furthermore the use of subsoil organic material is impacted by limited local availability, the high cost of suitable organic ameliorants delivered in-paddock, the sometimes large quantities required, the lack of suitable commercial-scale machinery and the poor predictability of when and where the amelioration will benefit crop productivity (Gill et al. 2008; Sale et al. 2019).
Gypsum application has been the most widespread traditional approach used to correct subsoil sodicity however the problems have included; surface application when the problem is evident in the subsoil, the large quantities of gypsum required to displace significant amounts of Na and the somewhat low solubility of gypsum.
Genetic improvement is also frequently advocated as an avenue for improving crop productivity and adaptation under different hostile soil conditions (McDonald et al. 2012; Nuttall et al. 2010). Little is known about genetic variation for subsoil constraints tolerance and how they relate to different plant traits such as elemental toxicity tolerance, canopy cover, rooting depth and harvest index and the integration of these factors in yield response of different genotypes. This limited knowledge is also due to the practical difficulties in measuring dynamic and variable soil constraints under field conditions.
To overcome sodic subsoil constraints within an economic framework a new approach was required that firstly commences with improved identification of the range of constraints present in the soil, the spatial variability of these constraints and synergising the combined approaches of inorganic and organic amendments with genetic solutions. This complex issue will be tackled by research within two GRDC investment projects (UA00159, DAV00149). This paper reports 2019 experimental results from these projects and is aimed at increasing WUE of grain crops on sodic subsoils by either ameliorating them with various organic and inorganic amendments or by identifying plant varieties and traits linked to sodicity tolerance.
Method
Rand amendment site
A field experiment was established on-farm near the township of Rand in southern NSW during February 2017. The site was located in a paddock that had been cropped with a cereal-canola rotation for more than 50 years. The physiochemical properties of the soil for this site are given in Figure 1. The soil is a Sodosol (Isbell 2002), with a texture-contrast profile increasing in clay content at depth. The increasing levels of exchangeable Na relative to calcium (Ca) and/or magnesium (Mg) in subsoil results in a decrease in soil structural stability and higher dispersion as shown in Figure 1. The high clay content in this subsoil layer has a bulk density of 1.55g/cm3 that restricts water movement, and consequently the saturated hydraulic conductivity value is low at 0.03cm/hr.
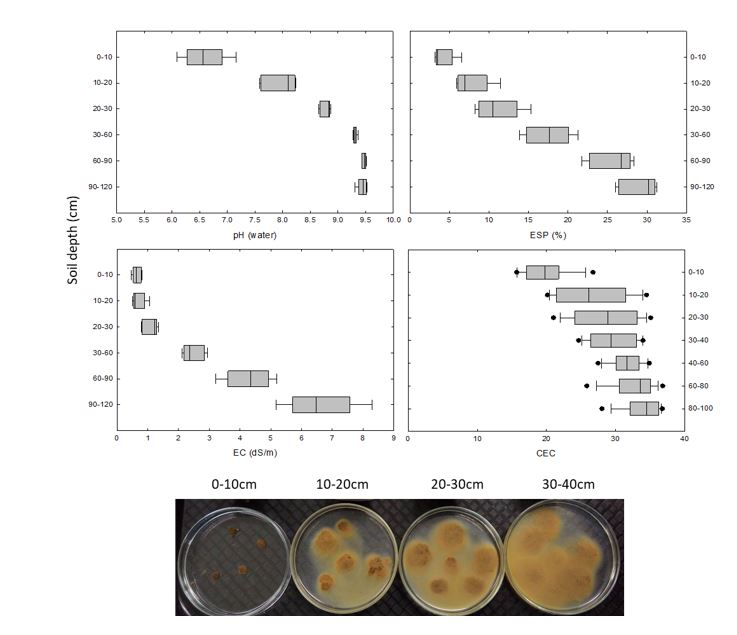
Figure 1. Soil characterisation of the Rand (southern NSW) experimental site. The picture shows the level of soil dispersion at four different depths.
The experimental plots were 2.5m wide and 20m long. There were 13 treatments comprising 1) the control, 2) surface application of gypsum, 3) surface application of chicken manure, 4) surface application of pea hay, 5) deep ripping, 6) deep placement of gypsum, 7) deep placement of chicken manure, 8) deep placement of wheat stubble, 9) deep placement of wheat stubble + nutrients, 10) deep placement of pea hay, 11) deep placement of pea hay + nutrients, 12) deep placement of liquid nutrients and 13) deep placement of pea hay + gypsum + nutrients. The experiment was a randomised complete block design with four replicates. Ripping and deep placement of amendments were carried out with a 3-D ripping machine (NSW DPI). The machine can deliver inorganic and/or organic amendments at two depths from 10cm to 30cm. The machine is also capable of delivering liquid fertilisers at depth. The experimental plots were sown to barley (cv. La Trobe) and wheat (cv. Lancer) in 2017 and 2018, respectively. In 2019 the experimental plots were sown on the 10 April to hybrid canola (cv. Pioneer®45Y92CL) at the seed rate of 4.4 kg/ha with an air seeder at 25cm row spacing. During sowing 90kg Mono-Ammonium Phosphate (MAP) (20kg P/ha and 9kg N/ha) was drilled in all plots and 100kg N/ha top-dressed in late June. In the 2019 growing season (April to November) this site received 215mm rainfall. Mean plant density as measured by seedlings counts three weeks after sowing was 37.7 ± 1.9 (mean ± SE of 60 plots) plants/m2.
At late flowering soil coring was completed using a tractor-mounted hydraulic soil-coring rig and 45mm diameter soil cores. The break core method was used to estimate rooting depth and exposed roots were recorded at the following depths 0 - 10, 10 - 20, 20 - 40, 40 - 60, and 60 - 100cm. Quadrat samples of 2m2 were taken at physiological maturity to measure plant biomass and grain yield.
Genotypes screening experiment
In 2019 an experiment was conducted at Grogan in southern NSW, which included 17 commercial wheat genotypes in a row column design with four replicates. The soil profile was slightly acidic in the top 10cm (pH1:5 water 5.9) and pH dramatically increased with depth (Table 1). The changes in soil sodicity (ESP) followed a similar trend as soil pH with ESP at 10.5% in the topsoil and increasing up to 40% in the subsoil (Table 1).
Table 1. Site characterisation for the Grogan experimental site. Values are means (n=5).
Soil depths (cm) | EC | pH (1:5 water) | Colwell-P | CEC (cmol(+)/kg) | Exchangeable sodium percentage |
|---|---|---|---|---|---|
0-10 | 309.40 | 5.87 | 58.80 | 16.66 | 10.53 |
10-20 | 133.00 | 7.65 | 7.40 | 22.06 | 11.97 |
20-30 | 136.90 | 8.76 | 2.62 | 24.53 | 15.94 |
30-40 | 207.66 | 9.12 | 2.50 | 25.55 | 20.12 |
40-60 | 338.94 | 9.60 | 1.34 | 27.17 | 26.27 |
60-80 | 530.40 | 9.53 | 1.00 | 31.63 | 36.68 |
80-100 | 897.20 | 9.43 | 1.48 | 34.07 | 40.25 |
100-120 | 1148.20 | 9.38 | 1.50 | 35.28 | 40.35 |
The experiment was sown on 17 May 2019 using a direct sown drill with DBS tynes spaced at 25cm. At sowing 90kg MAP (20kg P/ha and 9kg N/ha) was drilled in all plots and 75kg N/ha was surface applied just prior to stem elongation. Mean plant density, as measured by seedlings counts at four weeks after sowing, was 116 ± 1.6 (mean ± SE of 68 plots) plants/m2. At different growth stages multispectral images (MicaSense RedEdge-MX) were collected using drone technology to determine different vegetation indices such as normalised differences in vegetation index (NDVI) and leaf chlorophyll index (LCI) as a surrogate of canopy attributes and plant physiological processes (Liu et al. 2019; Satir and Berberoglu 2016; Zhang et al. 2019). Quadrat samples of 1m2 area were taken at physiological maturity to measure plant biomass and grain yield. Harvest index was calculated as grain yield divided by biomass.
Results
Rand amendment site
In 2019, canola grain yield significantly (P < 0.001) increased following the application of amendments in 2017 (Figure 2). The highest increase was observed for deep placement of pea hay + nutrient treatment and gypsum. Deep nutrients did not improve grain yield compared with the control and consequently it can be assumed the amendment affects were not due to nutritional factors only.
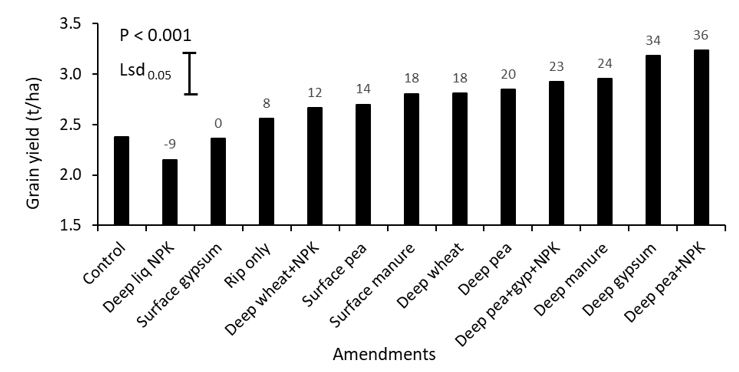
Figure 2. The mean effect of surface or deep-placed amendments on grain yield of canola (cv. Pioneer®45Y92CL) grown in alkaline sodic subsoil in Rand, southern NSW in 2019. Values on the top of each bar represent the percent change in grain yield for individual amendments compared to control.
The number of visible roots in the amended sodic subsoil (20 – 40cm depth) were also significantly (P < 0.05) affected by different amendments (Figure 3). Deep placement of both manure and pea hay increased the number of visible roots by more than three-fold. Neutron probe readings taken in September also indicate that the highest root counts were associated with the driest soil water profile (Figure 4). Variation in soil pH measured at different rooting depth during the flowering of canola is shown in Table 2. Compared to control, deep placement of gypsum reduced the soil pH by 0.7 units (8.8 to 8.1) at 20 – 40cm depth. However, pH was not affected by other treatments.
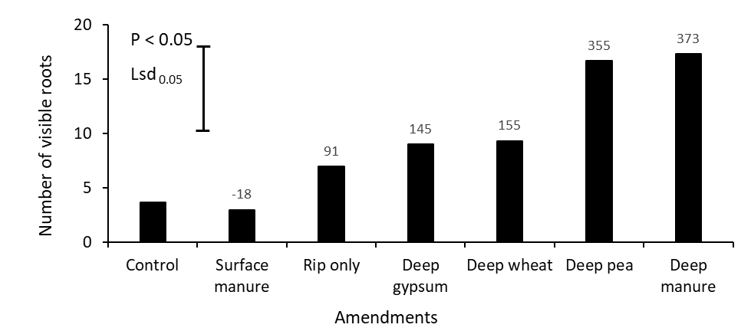
Figure 3. The mean effect of surface or deep-placed amendments on the number of visible roots at 30cm at late flowering of canola (cv. 45Y92CL) grown in alkaline sodic subsoil in Rand, SNSW in 2019. Values on the top of each bar is representing percent change of visible roots compared to control.
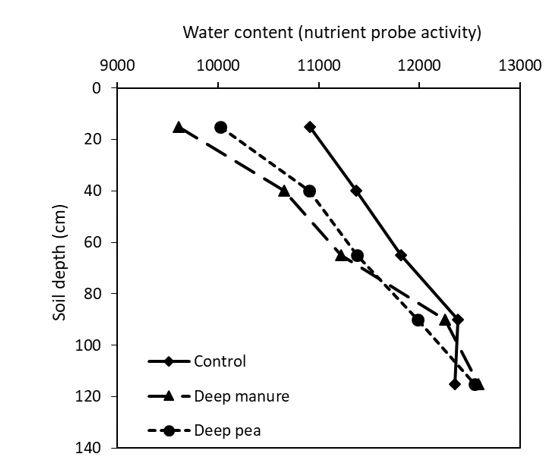
Figure 4. Neutron probe readings taken in September at the Rand amendment site for contrasting treatment comparisons. Results are based on the neutron activity (raw data) where higher values represent higher water content in the soil profile. Values are averages (n = 4).
Table 2. The changes in soil pH in selected treatments at Rand site. Samples were collected at late flowering of canola (cv. Pioneer® 45Y92CL) in September 2019.
Soil depths (cm) | Control | Surface manure | Rip only | Deep gypsum | Deep wheat | Deep pea | Deep manure |
|---|---|---|---|---|---|---|---|
0 - 10 | 6.5 | 6.8 | 6.7 | 6.9 | 7.3 | 7.1 | 6.8 |
10 - 20 | 7.7 | 7.8 | 8.1 | 7.1 | 8.1 | 8.2 | 8.0 |
20 - 40 | 8.8 | 8.7 | 8.8 | 8.1 | 8.9 | 9.0 | 8.5 |
40 - 60 | 9.3 | 9.4 | 9.3 | 9.3 | 9.4 | 9.5 | 9.5 |
60 - 100 | 9.5 | 9.3 | 9.4 | 9.3 | 9.4 | 9.6 | 9.3 |
Genotypes screening trial
Significant (P < 0.001) genotypic variation occurred in grain yield among the genotypes tested and ranged from only 0.57 t/ha (Gregory) to 2.0 t/ha (Scepter, Emu Rock and Mace; Figure 5). Biomass at final harvest did not significantly differ among the genotypes (data not shown; P = 0.11) and there was no significant (P = 0.09) correlation between grain yield and biomass at final harvest (Figure 6).
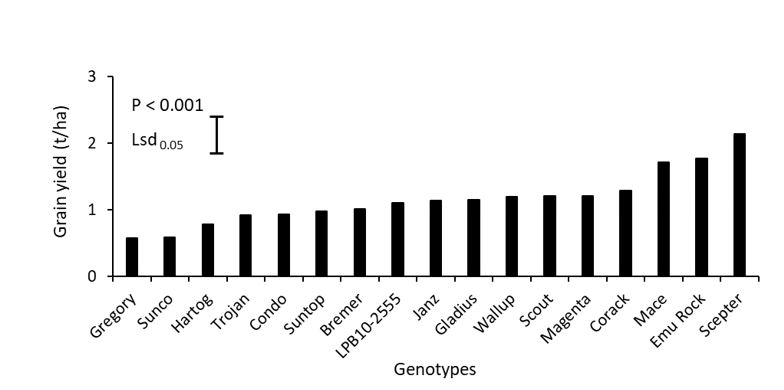
Figure 5. Variations in grain yield of 17 wheat genotypes grown in alkaline sodic dispersive subsoil in Grogan, southern NSW in 2019. Each data point is mean values of n = 4.
Significant variation was observed in the harvest index (data not shown; P < 0.001), which ranged from 0.08 (Gregory) to 0.26 (Scepter). A significant (P < 0.001) and positive correlation between harvest index and grain yield is observed among the studied genotypes (Figure 6).
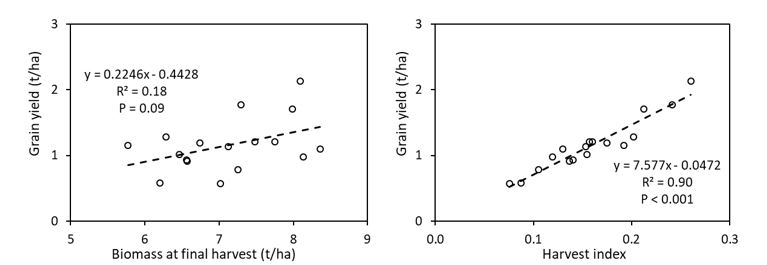
Figure 6. Linear regressions between grain yield and biomass at final harvest (left) and harvest index (right) of 17 wheat genotypes grown in alkaline sodic dispersive subsoil at Grogan, southern NSW in 2019.
All the non-destructive vegetation indices; NDVI (P < 0.01), normalised difference red edge (NDRE) (P < 0.001) and LCI (P < 0.001) measured at stem elongation showed significant and positive correlation with biomass at anthesis (Figure 7).
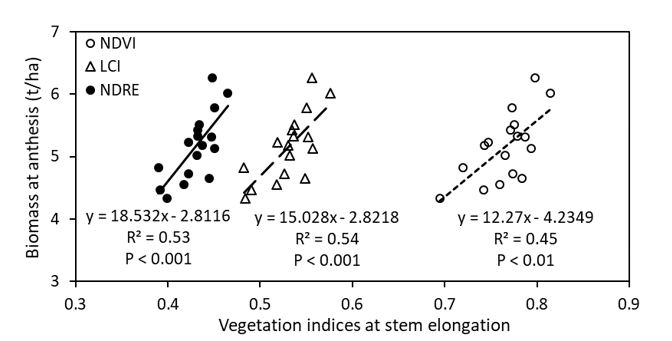
Figure 7. Linear regressions between vegetation indices (measured at stem-elongation) and anthesis biomass of 17 wheat genotypes grown in alkaline sodic dispersive subsoil at Grogan, southern NSW in 2019. NDVI = normalised differences in vegetation index; LCI = Leaf chlorophyll index; NDRE = Normalised difference red edge.
Discussion
The marked increase in the grain yield that occurred at Rand with the deep placement of both organic and inorganic amendments indicates the potential of this approach in reducing the yield gap associated with soil constraints in major cropping regions of Australia. Placement of the amendments in the study site in 2017 resulted in three consecutive years with significant yield improvement, indicating the residual effects of this approach on the yields of following crops (Gill et al. 2012). There was no positive yield response to deep nutrients and this supports other evidence that the responses at Rand are not due to nutritional factors alone. In a year of intensive drought like 2019, the grain yield improvements at Rand may be attributed to the additional root growth in the amended subsoil layer (Figure 3), which facilitated the use of extra subsoil water (Tavakkoli et al. 2019 and Figure 4). Under dryland conditions, water captured by roots in the subsoil layer is extremely valuable as its availability coincides with the grain filling period and has a very high conversion efficiency into grain yield (Kirkegaard et al. 2007; Wasson et al. 2012).
This study also indicates how the deep placement of both organic and inorganic amendments can improve soil chemicophysical properties. Reductions in extremely high soil pH and ESP at 20 – 40cm depth of the amended layers were reported within 14 months following deep placement of the amendments (Tavakkoli et al. 2019) and these changes have persisted into the third (2019) year (Table 2). Furthermore, improvement in soil chemical properties were also associated with increasing soil porosity, infiltration rate (data not shown) and microbial activity, which leads to soil aggregation and ultimately improving soil structure (Tavakkoli et al. 2019).
A major focus of this current research is to understand the amelioration processes of the subsoil application of organic and inorganic amendments. A tentative, but promising finding from our field and controlled environment trials is that farm-grown products like wheat and pea stubbles when mixed with nutrients improve soil aggregation, root growth, water extraction and grain yield and these treatments are comparable to animal manures and gypsum. If confirmed, this means that grain growers have a potentially large supply of relatively inexpensive organic ameliorants already available in their paddocks, which will increase the application options and viability of correcting subsoil sodicity.
Despite demonstrating significant improvements in grain yield with subsoil incorporation of organic and inorganic amendments, the widespread adoption of these practices is still limited by their cost effectiveness. Identifying traits associated with the superior tolerance to different soil constraints may be a low cost technique to tackle this issue (McDonald et al. 2012). Given the intensive drought condition in the study year, considerable genotypic variation was observed with some varieties having three- folds higher grain yield than the other varieties. Based on controlled-environment studies, the high yielding varieties at Grogan; Mace and Emu Rock, are moderately tolerant to tolerant to high pH and have roots that can grow relatively well through soils of high bulk density, whereas low yielding varieties such as Gregory, Hartog and Sunco are sensitive to one or both of these stresses. The very low harvest index in the trial suggests that there was severe stress around flowering to reduce grain set, as well as during grain filling and the results suggests that perhaps the ability to maintain root growth helped to alleviated the stress in varieties like Emu Rock and Mace. Furthermore, different traits associated with this greater yield performance of wheat genotypes are crucial aspects of future breeding programs.
Conclusions
The findings from the current field studies demonstrate initial but promising results of ameliorating alkaline sodic subsoils in medium and high rainfall zones of southern NSW. Deep placement of organic and inorganic amendments resulted in significant yield improvement in three successive years at Rand where subsoil water was present. This yield improvement was facilitated by a reduction in soil pH and ESP% and increased microbial activity that can lead to improved soil aggregation. Furthermore, deep placement of organic and inorganic amendments increased root growth, which in turn increased soil water use from the deeper clay layers during the critical reproductive stages of crop development, thereby increasing grain yield. In addition to soil management, genotypic variability in grain yield of wheat cultivars observed and their associated traits identified in the current study can be used for improving wheat germplasm through future breeding programs.
Acknowledgements
The research undertaken as part of this project is made possible by the significant contributions of growers through both trial cooperation and the support of the GRDC, the author would like to thank them for their continued support. This research was undertaken as part of projects; DAV00149 and UA000159. Thanks to Yan Jia (Soils Unit, NSW DPI) for technical inputs towards the NSW component of the project.
References
Adcock D, McNeill AM, McDonald GK, Armstrong RD (2007) Subsoil constraints to crop production on neutral and alkaline soils in south-eastern Australia: a review of current knowledge and management strategies. Aust J Exp Agric 47: 1245-1261. doi: 10.1071/ea06250.
Farooq M, Hussain M, Siddique KHM (2014) Drought stress in wheat during flowering and grain-filling periods. Critical Reviews in Plant Sciences 33: 331-349. doi: 10.1080/07352689.2014.875291.
Gill JS, Clark GJ, Sale PW, Peries RR, Tang C (2012) Deep placement of organic amendments in dense sodic subsoil increases summer fallow efficiency and the use of deep soil water by crops. Plant and Soil 359: 57-69. doi: 10.1007/s11104-012-1126-6.
Gill JS, Sale PWG, Tang C (2008) Amelioration of dense sodic subsoil using organic amendments increases wheat yield more than using gypsum in a high rainfall zone of southern Australia. Field Crops Research 107: 265-275. doi: 10.1016/j.fcr.2008.02.014.
Incerti M, O'Leary GJ (1990) Rooting depth of wheat in the Victorian Mallee. Aust J Exp Agric 30: 817-824. doi: 10.1071/ea9900817.
Isbell RF (2002) The Australian soil classification, Revised edn. CSIRO Publishing, Collingwood.
Kirkegaard JA, Lilley JM, Howe GN, Graham JM (2007) Impact of subsoil water use on wheat yield. Australian Journal of Agricultural Research 58: 303-315. doi: Crop and Pasture Science _ CSIRO Publishing.
Liu C, Liu Y, Lu Y, Liao Y, Nie J, Yuan X, Chen F (2019) Use of a leaf chlorophyll content index to improve the prediction of above-ground biomass and productivity. PeerJ 6: e6240-e6240. doi: 10.7717/peerj.6240.
McDonald GK, Taylor JD, Verbyla A, Kuchel H (2012) Assessing the importance of subsoil constraints to yield of wheat and its implications for yield improvement. Crop & Pasture Science 63: 1043-1065. doi: 10.1071/cp12244.
Nuttall JG, Hobson KB, Materne M, Moody DB, Munns R, Armstrong RD (2010) Use of genetic tolerance in grain crops to overcome subsoil constraints in alkaline cropping soils. Australian Journal of Soil Research 48: 188-199. doi: 10.1071/sr09081.
Orton TG, Mallawaarachchi T, Pringle M, Menzies NW, Dalal RC, Kopittke PM, Searle R, Hochman Z, Dang YP (2018) Quantifying the economic impact of soil constraints on Australian agriculture: A case-study of wheat. Land Degradation & Development 29: 3866-3875. doi: 10.1002/ldr.3130.
Passioura JB, Angus JF (2010) Improving productivity of crops in water-limited environments. In: DL Sparks (ed) Advances in Agronomy, Vol 106.
Sale PW, Gill JS, Peries RR, Tang C (2019) Crop responses to subsoil manuring. I. Results in south-western Victoria from 2009 to 2012. Crop and Pasture Science 70: 44-54. doi: Crop and Pasture Publishing _ CSIRO Publishing.
Satir O, Berberoglu S (2016) Crop yield prediction under soil salinity using satellite derived vegetation indices. Field Crops Research 192: 134-143. doi: Crop yield prediction under soil salinity using satellite derived vegetation indices - Science Direct
.
Tavakkoli E, Weng ZH, Tahmasbian I, Uddin S, Poile G, Oates A, Xu BB, Sandral G, Fang Y, Armstrong R (2019) Understanding the amelioration processes of the subsoil application of amendments. 18 - 19 February 2019, GRDC Grains Research Update (Wagga Wagga).
Wasson AP, Richards RA, Chatrath R, Misra SC, Prasad SVS, Rebetzke GJ, Kirkegaard JA, Christopher J, Watt M (2012) Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops. J Exp Bot 63: 3485-3498. doi: 10.1093/jxb/ers111.
Zhang K, Ge X, Shen P, Li W, Liu X, Cao Q, Zhu Y, Cao W, Tian Y (2019) Predicting rice grain yield based on dynamic changes in vegetation indexes during early to mid-growth stages. Remote Sensing 11: 387.
Contact details
Dr Ehsan
NSW DPI, Wagga Wagga Agricultural Institute
02 69381992
Ehsan.tavakkoli@dpi.nsw.gov.au
@EhsanTavakkoli
Dr Shihab Uddin
Shihab.uddin@dpi.nsw.gov.au
Varieties displaying this symbol beside them are protected under the Plant Breeders Rights Act 1994.
® Registered trademark