Soil organic matter in dryland systems – management and opportunities
Soil organic matter in dryland systems – management and opportunities
Take home messages
- It is well known that stocks of soil organic carbon have declined over the past decades in many Australian agricultural systems, including dryland grains production.
- This loss of carbon (C) has also resulted in a substantial reduction in soil nutrient stocks, particularly nitrogen (N), that supply a large proportion of a crop’s nutrition.
- Rebuilding soil C and N stocks is slow and depends upon increased biomass production to drive C inputs, supported by fertiliser requirements being met. It is possible this may be assisted by some regenerative approaches.
- Identifying and addressing the primary constraints to water use efficiency, inclusion of legumes in crop sequences, and estimating and managing the N budget of the system over multiple seasons should turn a system towards rebuilding C and N stocks.
- Practices that fall under the umbrella definition of regenerative agriculture may limit C losses, or assist in building stocks, provided other constraints are overcome.
- This paper is largely a revision of Farrell et al. (2021) and Macdonald et al. (2020).
Introduction
Soil organic matter (SOM) contains the largest stocks of both C and N, including those under agricultural management. Globally, 1200—1550 gigatonnes (Gt) of C are stored in soils, with estimates of 22.6–39.7Gt in the top 30cm of Australian soils (Viscarra Rossel et al. 2014). Assuming a C:N ratio of 11.8, this equates to 1.92–3.36Gt N stored in the SOM of the top 30cm of all of Australia’s soils, an average of just over 4t N/ha. Most N is not immediately available, and is bound within SOM as organic N. As plants can only take up mineral N and a small proportion of dissolved organic N (DON), SOM must be decomposed to release these compounds. A snapshot study found, on average, only 0.59—4.80% of total N was present in plant available forms in Australian agricultural systems (Farrell et al. 2016).
Soil organic matter is responsible for provision of nutrients (particularly N), maintaining a diverse and healthy microbial community, infiltration and water retention, amongst others. It can be separated into three measurable fractions, with distinct properties for nutrient supply and stability (Baldock et al. 2013), which were described in detail in our update to GRDC last year (Farrell et al. 2021). In addition to SOC, two smaller important pools of C exist: microbial biomass C (MBC), which typically contains approximately 1% of the total organic C in a soil and represents the C stored in live microorganisms, and dissolved organic C (DOC) which is the soluble fraction. This latter pool contains most of the C directly accessible by microorganisms, but also compounds that, while soluble, do not directly reflect availability of C. Though representing only a small percentage of C in the soil, they turn over quickly and yet are very important. In many ways, this could be thought of in terms of a bath: If you were to run the tap with the plug out and measure the amount of water in the bath, you would find very little there at any point in time. However, you would be wrong to conclude that water is not important for the function of a bath. This same concept of ‘flux’ versus the ‘pool’ needs to be considered for N also.
Regenerative agriculture practices
As many would be aware, there has been a groundswell in the ‘regenerative agriculture movement’ over the past few years. This primarily comes from overseas, and at its simplest, aspires to effectively leaving the land in a better condition than that in which it started. Robertson et al. (2022) propose an elaborated definition of regenerative agriculture (RA):‘Regenerative agriculture is a form of farming in which explicit attention is paid to the state and trajectory of the natural capital base (soil, water, biodiversity) underlying farm production and acknowledgement that there are non-farm stakeholders interested in its responsible management. It is not a prescriptive recipe of farm practices, but rather has a focus on positive outcomes for the natural resource base, particularly soil health, and farm productivity and profitability.’
Whilst not prescriptive (unlike organic or biodynamic systems), typically promoted RA practices include:
- minimum soil disturbance
- stubble retention
- diverse rotations (including cover crops and the inclusion of livestock in the system)
- a reduction in synthetic inputs (including pesticides and fertilisers).
In Australian broadacre agriculture, minimum soil disturbance, through the adoption of no-till (NT) and stubble retention, have been almost universally adopted over the past 20-30 years, whilst diverse rotations are increasingly seen as a best practice way to manage disease and the risk of seasonal variations. Thus, the first three RA practices, far from being something new, are widely adopted conventional farming practices in Australia, with the general presumption that amongst other agronomic benefits, SOC stocks also increase. Contrasting findings in the literature, even between global meta-analyses (Kopittke et al. 2017; Powlson et al. 2014) suggest this outcome to be variable, and likely climate- and soil- specific. Sanderman et al. (2010) reported improved cropping practices in Australia has the potential to increase SOC stocks by 0-2-0.3t/ha/yr, though many of the improvements within that definition (for example, NT, stubble retention, and diverse rotations) are now well established as best practice.
On the other hand, in many cases, reducing the amount of synthetic inputs is a far more complex topic, separable into two distinct categories:
- agrichemicals for weed, pest and disease control
- synthetic fertilisers.
In-depth discussion of the impact of agrichemicals is beyond the scope of this paper, but it is well known that NT systems rely primarily upon chemical methods for weed control. Conversely, negative impacts of mechanical weed control via regular tillage on soil structure and erosion are well documented.
When considering an aspiration to reduce synthetic fertiliser inputs, particularly N, it is important to recognise the intrinsic links between SOC and N — the majority of N in soils is chemically bound to SOC in the form of SOM. If synthetic N inputs are reduced and the balance is not replaced via N fixation, N will be ‘mined’ by the crop from SOM, resulting in a loss of SOC. This is discussed further below.
Linkages between the soil carbon and nitrogen cycles
Organic matter – form and function
To better understand how C accumulates, is lost and behaves in soil, a fractionation procedure has been developed to separate measurable fractions of discrete chemistry and functionality. This separates C in the <2 mm portion of soil (excluding gravel and coarse plant debris) into three fractions:
- Particulate organic C (POC): The least decomposed fraction that is accumulated rapidly but also is most vulnerable to loss and is dominated by partially decomposed plant material.
- Humus-like organic C (HOC): Stabilised organic carbon, mostly in the form of dead soil microorganisms, that has undergone degradation and is often protected from loss due to binding to the soil mineral phase and protection within microaggregates.
- Resistant organic C (ROC): This is a charcoal-like substance, typically with a very high C:N ratio >100:1 as SOM and a residence time of millennia.
Inputs and retention of C in dryland soils
In Australian broadacre cropping systems, there are typically only two sources of C input to soil: the C fixed by plants in the paddock through photosynthesis, and the C contained in organic amendments such as manures and composts that may be applied. Though encouraged, if available at a reasonable price close to a source, the import of organic matter is not a viable option in broadacre agriculture in many locations. Thus, the focus of this section is primarily on inputs from the crop or pasture plants grown in situ.
As discussed in more detail in Macdonald et al. (2020) and Farrell et al. (2021), it is possible to estimate the inputs of plant C to the soil on the basis of observed crop yield. This can be done on the basis of several literature-derived figures for the key aspects of harvest index, C allocation within a crop, and retention factors for C additions to soil (Figure 1).
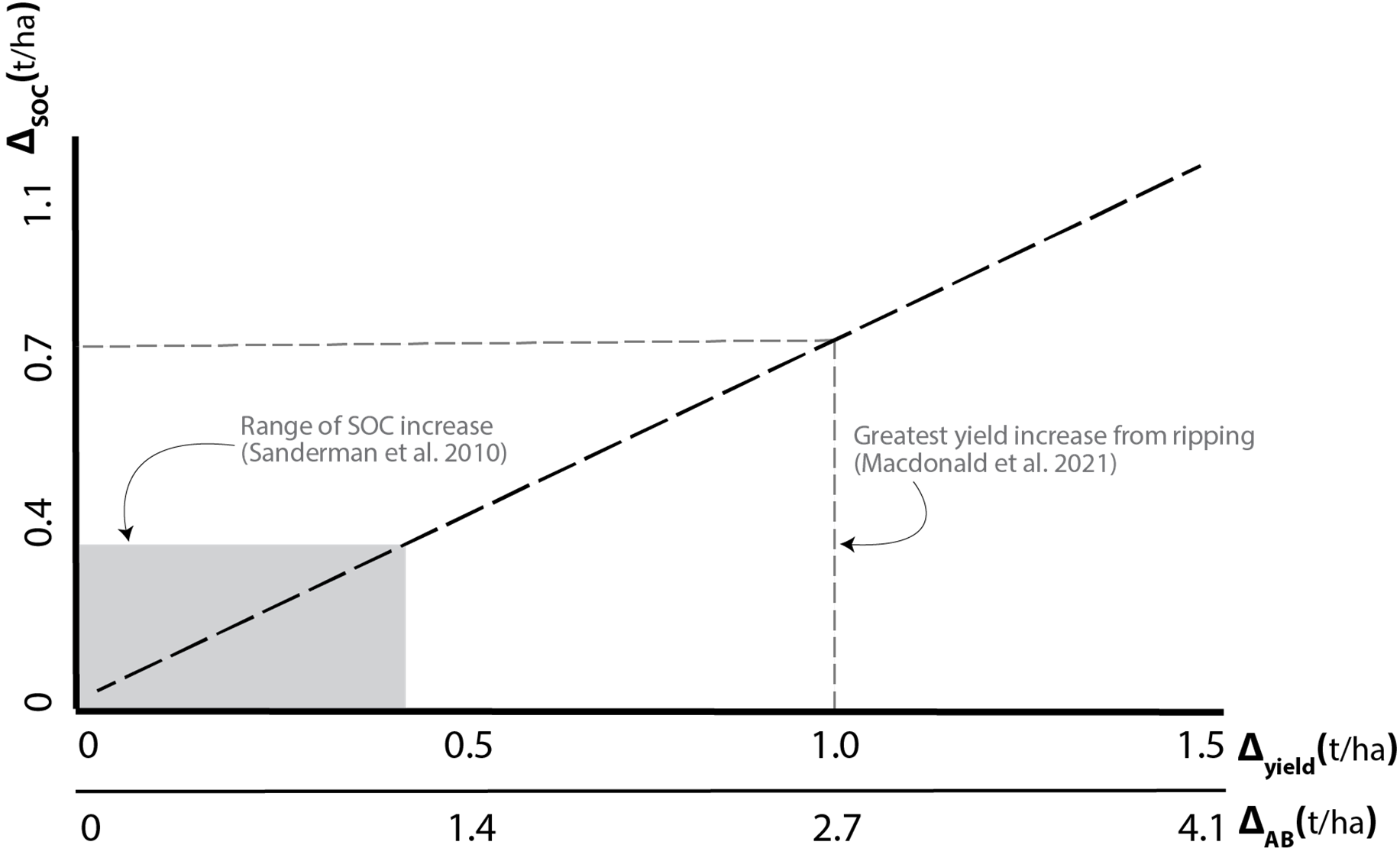
Figure 1. Relationship between changes in grain yield ( ; 1st x-axis) or aboveground biomass ( ; 2nd x-axis) and changes in SOC ( ; y-axis) calculated using equations and data presented in Farrell et al. (2021). Values are estimates and per year. The greyed area shows the range for the published studies summarised by Sanderman et al. (2010) where ‘improved management practices’ resulted in an increase of up to 0.3t SOC/yr, a probable yield increase of approximately 0.4t/ha grain would be required. The grey dashed lines show the estimated change in SOC as a result of the average greatest yield increase realised as a result of deep ripping and associated activities in the current GRDC ‘Sandy Soils’ project (Macdonald et al., 2021).
It is important to note that many of the factors used in the derivation of Figure 1 would be expected to vary in a non-linear manner with yield, and thus increases in yield in the higher ranges may not result in the same proportion of photosynthetically-fixed C being translocated to the non-grain pools. Further, retention factors of C in the soil are likely to be very soil-type dependent, and it is unlikely that such figures would apply equally across different soil classes and textures, with higher retention likely in heavier clay soils. Lastly, these figures do not consider losses of existing C, either through priming (Chowdhury et al. 2014) or as a result of disturbance in more energy-intensive amelioration activities, such as the deep ripping reported in Macdonald et al. (2021).
Without nitrogen, there is little opportunity to increase carbon
The main input of N in broadacre cropping comes either from fertiliser or the inclusion of legumes in a rotation sequence, although a small proportion may also arrive through atmospheric deposition and fixation by free-living microbes in the soil. It is generally perceived that the efficiency of fertiliser N in Australian grains systems is low, with 40-50% of the N applied to a crop being recovered in that crop within the same season (Angus and Grace 2017). That does not however mean that the remaining N is lost from the system. In Australian dryland cropping systems, losses of fertiliser N through leaching are low, especially in low rainfall zones. Further, while gaseous emissions of N2 and NH3 are less well quantified and may contribute substantial losses of N in some situations (Harris et al. 2016), N2O emissions are amongst the lowest of any managed agricultural system.
Even in more heavily fertilised irrigated cotton systems of New South Wales and Queensland, the majority of N taken up by a crop is accessed via soil processes; primarily SOM mineralisation (Macdonald et al. 2016), and thus, the acknowledgement that efficiency of fertiliser N use sits at approximately 50% in a given season obscures the use of N supplied in previous seasons.
The ‘elephant in the room’ when it comes to N export or loss from farming systems is actually the amount that is removed in the produce itself. As reported at previous GRDC Updates events, a comprehensive study by Norton (2016) found that the majority of properties studied in southern and eastern Australia were net exporters of N from the crop alone. Harries et al. (2021) found negative N balances in 60% of paddocks surveyed in WA. If other losses (particularly N2 from denitrification, which is the least quantified) are also present in the system, this could contribute to substantial N mining in the medium to longer term, with concomitant impacts on SOC stocks (Baldock et al. 2018).
Management to build carbon and nitrogen stocks
For both C and N, the same principle applies; the stock in the soil is a function of inputs and outputs. While the inputs of N are perhaps somewhat simpler to conceptualise and manage, it is primarily plant growth that results in C inputs to soil, and there are several means of manipulating this to improve the likelihood of increasing C stocks. Also, as C stocks increase over the longer term, it is likely that the ability of the soil to supply N through the mineralisation of SOM will also increase, provided that the N balance remains positive.
Addressing soil constraints to increase soil organic matter inputs
Despite gains in productivity from broad adoption of NT and early sowing, there remains a yield gap between paddock production and the water limited yield potential. In many Australian cropping systems, crop water use is limited by a range of surface and subsurface constraints which limit root growth and exploration. Common abiotic constraints include compaction, soil acidity and associated toxicities (aluminium, magnesium), alkalinity, sodicity and associated toxicity (boron, chloride, salt), and water repellence. Biotic constraints are also recognised as important in Australian agriculture and include disease, weed, and pest pressures (Lawes et al. 2021). While conservation practices and crop selection are useful tools in mitigating the impact of these constraints, they will not correct the physico-chemical condition of the soil.
Amelioration practices aim to overcome soil constraints for long-term improvement to crop growth and productivity. Under these scenarios, crop productivity and biomass production can be greatly increased and will have subsequent impact on C and N flows through the soil profile. Examples include current research targeting subsoil acidity (Fleming et al. 2020), and deep ripping combined with the addition of organic amendments resulting in yield gains in some situations between 0.4 – 2t/ha (Macdonald et al. 2021). Research is ongoing to assess the longer-term benefits of these approaches to manage soil constraints, including developing a clearer understanding of scenarios in which they can be relied upon to deliver clear cumulative yield increases.
Nitrogen balance, excess and the ‘nitrogen bank’
Typical N fertiliser decisions focus either on rules of thumb for a district, or predictions of yield, and thus, likely N demand on the basis of model predictions, for example, Yield Prophet®. By and large, such predictions target maximum profit over yield. Importantly, they typically focus on returns within a single season. As Norton (2016) has shown, the net N balance of such practice is usually negative, meaning that N is being ‘mined’ from the SOM at a greater rate than it is returned, resulting in a concomitant loss of C. Thus, yields are effectively being ‘subsidised’ by SOM loss resulting in medium-long term reduction in the ability of soil to supply N through in-season mineralisation. This reduces the soil’s fertility in the longer term, and as mineralisation tends to release N at a rate closely matching the crop’s pattern of N demand, it is unlikely that extra fertiliser can simply offset lost N mineralisation potential in the longer term. Further, the main factor driving the yield gap in Australian grains systems is N limitation (Hochman and Horan 2018), and in wet and favourable seasons, this conservative approach to N management may impact profit in the short-term.
Instead, growers and advisers could consider the N requirement of the system as a whole by:
- considering nutrient balance in the medium-long term, namely N balance over a 5–10-year period, not just the season ahead, and
- considering the need to ‘feed the soil’ via immobilisation of nutrients, as much as the crop itself.
This second point explicitly accounts for the fertiliser N required to build SOM which is sometimes seen as a negative cost of building C stocks but allows for the replenishment of the store of N that is released slowly through mineralisation.
An emerging approach to slow and potentially reverse declines in SOC and N stocks is known as the ‘nitrogen bank’ strategy (Meier et al. 2021). Recognising that losses of N from dryland grains systems are often low, and thus, economic and environmental risks are minimal (Smith et al. 2019), we suggest that applying greater rates of N will increase profitability through addressing the main constraint to yield and reducing SOC run-down. A major limitation in calculating a crop’s N requirement is the ability to forecast rainfall and water-limited yield potential early in the season. A simple solution to this uncertainty is proposed whereby fertiliser application is calculated as the balance of crop N demand required to achieve economic yield in the majority of seasons after subtraction of the available N stock at sowing, ignoring in-season mineralisation. If it is a dry season, excess N will mostly remain in situ and be captured in the next season’s pre-sowing N testing, and fertiliser application rates adjusted accordingly. This approach effectively removes much reliance on SOM to deliver N through in-season mineralisation, and SOM that is mineralised is likely replaced through greater plant C inputs and the higher N availability resulting from the increased fertilisation rates. It should be noted that whilst showing early promise with minimal fertiliser N losses in the drier systems that dominate the southern region (Smith et al. 2019), substantial losses through denitrification of larger up-front N additions have been documented in ex-pasture systems in the high rainfall zone (for example, up to ~90% applied N, Harris et al. 2016). Further research is required to better understand the climatic and soil boundaries at which higher up-front N applications can be applied with minimal loss.
Regardless of the system or strategy (Yield Prophet, N banks, rules of thumb) used to decide how much N to apply, a long-term field experiment in the Victorian Mallee has shown that systems which run a neutral to small positive N balance (that is, are not mining SOM) are also the most profitable (Figure 2).
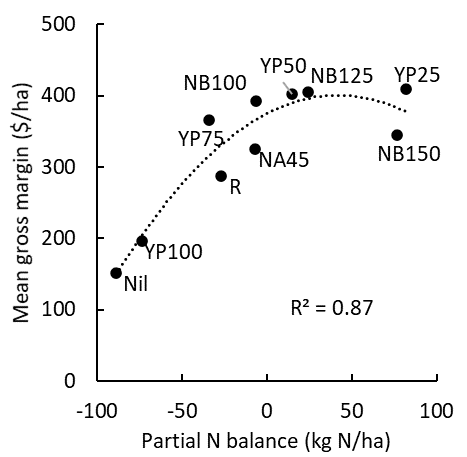
Figure 2. The relationship between partial N balance (fertiliser N additions minus offtake in grain) and mean gross margin from a 4-year BCG experiment (2018-2021) at Curyo in NW Victoria. NB=nitrogen bank at different targets (100, 125 and 150kg N/ha); YP=Yield Prophet® at different levels of probability (25, 50, 75 and 100% where 50% targets median seasonal yield potential); R=replacement (N offtake in grain balanced with fertiliser); NA=National Average (45kg N/ha). For further details see Birchip Cropping Group.
Legumes and nitrogen fixation
One of the key benefits of grain and pasture legumes in crop rotation is the N contribution through legume-rhizobia symbiosis that provides a component of the legume N requirements and is an important contributor of N supply to subsequent crops. The effect of recent intensification of Australian cropping systems and the consideration of grain legumes as rotational crops has the potential to reduce N inputs and increase N use efficiency in following crops and improve overall soil quality. It is generally accepted that for many legume species, on average 20kg of shoot-N per tonne of dry matter is fixed by grain legumes, although the actual amount of N fixed can vary 15 — 25kg N fixed per tonne depending on the legume type, field conditions including management practices applied and seasonal conditions (Peoples et al. 2009). However, it should be remembered that N fixation provides the majority of the N demand of the grain legume crop itself, and a large part of the fixed N is exported in the grain.
Despite the increased preference for cropping in recent years, pastures remain a dominant part of southern farming systems which can play a key role in sustaining and improving SOM and fertility. Nitrogen fixation from pastures provides an important component of the N supply to subsequent cereal crops, which are further complemented by the C inputs from above and below ground plant components. The amount of N fixed by various annual and perennial legumes in Australia can vary from <10 to >250kg N/ha/year. Additionally, the below-ground pool of N in roots and nodules provides an important source of N inputs, for example, 40-55% of total plant N is estimated to be present below ground in pasture systems (Peoples et al. 2017).
There is an opportunity to improve our understanding of the constraints to N fixation. For example, inappropriate herbicide use has been shown to reduce N fixation in grain legumes (Drew et al. 2007). Implementation of management strategies that can improve legume productivity and N fixation can not only arrest the decline in the N supply capacity of soils but also contribute to the improvement of overall SOM quantity and quality (Sanderman et al. 2017). Given that the formation of new SOM is not only contingent on there being sufficient C and nutrients, but also the need for them to be co-located near clay minerals and in conditions suitable for microbial growth, conversion of legume root biomass to more stabilised SOM is likely to be more efficient than other plant inputs supplemented with nutrients supplied by fertiliser.
Cover crops
A final potential strategy to build C stocks, improve soil resilience and address N decline is the implementation of break or cover-cropping, either as green/brown manure or to provide supplemental stock feed. Winter cover crops may be grown in lieu of a cash crop as part of a rotation sequence, or they may be established opportunistically during the summer fallow. With regards to managing the soil, their aim is to reduce erosion by maintaining a ground cover, increase C inputs and microbial activity, reduce soilborne disease impacts and potentially address nutrient stratification or subsoil constraints through deep roots.
A recent study in Europe found that the length of vegetation cover was more important for grain yields and soil function than diversity within a rotation (Garland et al. 2021). If sown as a species mixture, the combination of species can be tailored to occupy multiple niches so that biomass production ‘overyields’, that is, produces more biomass than that from an equivalent monoculture.
Cover cropping is an increasingly adopted strategy overseas, particularly in the USA. However, in Australia’s much drier climate, substantial questions remain as to whether any benefits derived offset potential loss of water through evapotranspiration of the cover crop, particularly in summer applications where the prevailing guidance is to manage weeds to maximise soil water retention. Current research led by Agex, SANTFA and CSIRO (Farrell and Stanley, 2021) is exploring these issues across 20 sites in the southern region, and is due to report later this year.
Looking to the future
A growing body of evidence suggests that fertiliser strategies designed to maximise profit or offset financial risk in the short term do not meet the N demand of the system, and thus, invoke N-mining and resultant SOC loss. To arrest and reverse the loss of C and concomitant draw-down of N reserves in soils, the simple equation is that inputs need to be greater than exports and losses. There are several ‘levers that can be pulled’ on both sides of this equation, but it is important to understand that for the most part, the soil C and N cycles are intrinsically linked, as most N is bound in SOM, and the effectiveness of management efforts will be strongly influenced by climate and soil type (Hunt et al.2020). Approaches that increase N inputs will both reduce N-mining and increase C inputs through greater plant productivity.
Recognising the monetary value of the N tied up in SOM (and indeed, exploited through N-mining) suggests that a longer-term approach to N fertilisation strategies and legume rotations which result in a net import of N are required. The nitrogen bank strategy provides one possible solution whereby longer-term profitability and higher N applications are not mutually exclusive. Coupled with strategies that increase plant C inputs, either through the alleviation of soil constraints or where appropriate, increased plant growth and time of soil cover through cover cropping, it is likely that, over time, SOM and thus, N and C stocks, will increase. It is possible to achieve a neutral or positive N balance with RA and thus build SOM, but this likely requires 50% of rotation in pasture legumes or winter cover crops, or affordable access to organic inputs to balance N removed in grain. In some regions with good access to waste streams, this may be feasible, whilst in other regions it would be cost prohibitive.
Many growers and advisers will ask ‘Why should we do this? Can we offset the rundown of soil N over the longer term by just increasing fertiliser rates once yields drop?’ The pragmatic answer is perhaps ‘maybe…’. However, mineralisation of SOM mimics N demand of crops and this is difficult to match with fertilisers, even advanced slow-release formulations. It seems highly unlikely that increased reliance on fertiliser N will improve the efficiency of N use by crops at the system level, with increased losses and lower efficiency of use in the longer term. Of course, the delivery of N is but one of the many ecosystem services we rely upon SOM to deliver. Rundown of soil N and associated loss of SOM would also result in reduced microbial activity and likely capacity to buffer against disease impacts.
References
Angus JF, Grace PR (2017) Nitrogen balance in Australia and nitrogen use efficiency on Australian farms. Soil Research55(6), 435–450. doi:10.1071/SR16325
Baldock JA, Farrell M, Macdonald LM, Welti N, Monjardino M (2018) ‘Nutrient mineralisation in Australian grain producing soils: mechanisms, magnitudes, gaps and future research.’ (CSIRO: Canberra).
Baldock JA, Sanderman J, Macdonald LM, Puccini A, Hawke B, Szarvas S, McGowan J (2013) Quantifying the allocation of soil organic carbon to biologically significant fractions. Soil Research51(8), 561-576.
Chowdhury S, Farrell M, Bolan N (2014) Priming of soil organic carbon by malic acid addition is differentially affected by nutrient availability. Soil Biology and Biochemistry 77, 158–169. doi:10.1016/j.soilbio.2014.06.027
Drew EA, Gupta VVSR, Roget DK (2007) Herbicide use, productivity, and nitrogen fixation in field pea (Pisum sativum). Australian Journal of Agricultural Research 58(12), 1204-1214.
Farrell M, Allen DE, Macdonald BCT (2016) Pools and fluxes: a snapshot of nitrogen dynamics in Australian soils. Proceedings International Nitrogen Initiative Conference, Melbourne, December 2016, pp. 1–4.
Farrell M, Gupta V, Macdonald LM (2021) Addressing the rundown of nitrogen and soil organic carbon. Proceedings GRDC Grains Research Update, Bendigo, February 2021, pp. 51-60.
Farrell M, Stanley M (2021) Mixed cover crops for sustainable farming’ (CSIRO)
Fleming N, Fraser M, Dohle L, Hughes B (2020) Subsurface acidity – how far has the research advanced? Proceedings GRDC Grains Research Update, Adelaide, February 2020, pp. 251-256.
Garland G, Edlinger A, Banerjee S, Degrune F, Garcia-Palacios P, Pescador DS, Herzog C, Romdhane S, Saghai A, Spor A, Wagg C, Hallin S, Maestre FT, Philippot L, Rillig MC, van der Heijden MGA (2021) Crop cover is more important than rotational diversity for soil multifunctionality and cereal yields in European cropping systems. Nature Food 2, 28–37. doi:10.1038/s43016-020-00210-8
Harries M, Flower KC, Scanlan CA (2021) Sustainability of nutrient management in grain production systems of south-west Australia. Crop and Pasture Science 72(3), 197-212.
Harris RH, Armstrong RD, Wallace AJ, Belyaeva ON (2016) Effect of nitrogen fertiliser management on soil mineral nitrogen, nitrous oxide losses, yield and nitrogen uptake of wheat growing in waterlogging-prone soils of south-eastern Australia. Soil Research 54(5), 619-633.
Hochman Z, Horan H (2018) Causes of wheat yield gaps and opportunities to advance the water-limited yield frontier in Australia. Field Crops Research 228, 20–30. doi:10.1016/j.fcr.2018.08.023
Hunt JR, Celestina C, Kirkegaard JA (2020) The realities of climate change, conservation agriculture and soil carbon sequestration. Global Change Biology 26(6), 3188–3189. doi:10.1111/gcb.15082
Kopittke PM, Dalal RC, Finn D, Menzies NW (2017) Global changes in soil stocks of carbon, nitrogen, phosphorus, and sulphur as influenced by long‐term agricultural production. Global Change Biology 23(6), 2509–2519. doi:10.1111/gcb.13513
Lawes R, Chen C, Whish J, Meier E, Ouzman J, Gobbett D, Vadakattu G, Ota N, van Rees H (2021) Applying more nitrogen is not always sufficient to address dryland wheat yield gaps in Australia. Field Crops Research 262, 108033.
Macdonald BCT, Chang YF, Nadelko A, Tuomi S, Glover M (2016) Tracking fertiliser and soil nitrogen in irrigated cotton: uptake, losses and the soil N stock. Soil Research 55(3), 264–272. doi:10.1071/SR16167
Macdonald LM, Farrell M, Baldock JA (2020) Soil organic matter – what the science tells us. Proceedings GRDC Grains Research Update, Barellan, March 2020, pp. 28–35.
Macdonald LM, Wilhelm N, Fraser M, Moodie M, Trengove S, Desbiolles J, Saunders C, Whitworth R, da Silva R, Llewellyn R, McBeath T (2021) Targeted amelioration in Mallee sands to maximise crop water use. Proceedings GRDC Grains Research Update, Adelaide, February 2021, pp. 37-42.
Meier EA, Hunt JR, Hochman Z (2021) Evaluation of nitrogen bank, a soil nitrogen management strategy for sustainably closing wheat yield gaps. Field Crops Research 261, 108017. doi:10.1016/j.fcr.2020.108017
Norton RM (2016) ‘Nutrient performance indicators from southern Australian grain farms.’ (GRDC, Canberra).
Peoples MB, Brockwell J, Herridge DF, Rochester IJ, Alves BJR, Urquiaga S, Boddey RM, Dakora FD, Bhattarai S, Maskey SL, Sampet C, Rerkasem B, Khan DF, Hauggaard-Nielsen H, Jensen ES (2009) The contributions of nitrogen-fixing crop legumes to the productivity of agricultural systems. Symbiosis 48(1-3), 1–17.
Peoples MB, Swan AD, Goward L, Kirkegaard JA, Hunt JR, Li GD, Schwenke GD, Herridge DF, Moodie M, Wilhelm N, Potter T, Denton MD, Browne C, Phillips LA, Khan DF (2017) Soil mineral nitrogen benefits derived from legumes and comparisons of the apparent recovery of legume or fertiliser nitrogen by wheat. Soil Research 55(6), 600–616. doi:10.1071/SR16330
Powlson DS, Stirling CM, Jat ML, Gerard BG, Palm CA, Sanchez PA, Cassman KG (2014) Limited potential of no-till agriculture for climate change mitigation. Nature Climate Change 4, 678–683. doi:10.1038/nclimate2292
Robertson M, Macdonald B, Farrell M, Norman H, Macdonald L, Vadakattu G, Taylor J (2022) What can science offer the proponents of Regenerative Agriculture practices? Farm Policy Journal, accepted.
Sanderman J, Creamer C, Baisden WT, Farrell M, Fallon S (2017) Greater soil carbon stocks and faster turnover rates with increasing agricultural productivity. Soil 3, 1–16. doi:10.5194/soil-3-1-2017
Sanderman J, Farquharson R, Baldock JA (2010) Soil carbon sequestration potential: a review for Australian agriculture. CSIRO Sustainable Agriculture Flagship, Adelaide, Australia.
Smith CJ, Hunt JR, Wang E, Macdonald BCT, Xing H, Denmead OT, Zeglin S, Zhao Z (2019) Using fertiliser to maintain soil inorganic nitrogen can increase dryland wheat yield with little environmental cost. Agriculture Ecosystems & Environment 286, 106644. doi:10.1016/j.agee.2019.106644
Viscarra Rossel RA, Webster R, Bui EN, Baldock JA (2014) Baseline map of organic carbon in Australian soil to support national carbon accounting and monitoring under climate change. Global Change Biology 20(9), 2953–2970. doi:10.1111/gcb.12569
Contact details Mark Farrell
CSIRO Agriculture & Food
Locked Bag 2, Glen Osmond SA 5064
08 8303 8664
mark.farrell@csiro.au
@inverted_soil