Recent improvements in biomass partitioning and transpiration efficiency of modern Australian wheat varieties – any opportunity for the future?
Recent improvements in biomass partitioning and transpiration efficiency of modern Australian wheat varieties – any opportunity for the future?
Author: Andrew Fletcher and Karine Chenu, University of Queensland, Queensland Alliance for Agriculture and Food Innovation (QAAFI) | Date: 01 Mar 2016
Take home message
- By selecting for greater yield in rainfed conditions, Australian wheat breeders have historically selected indirectly for varieties with greater transpiration efficiency, which produce more crop per drop.
- Modern wheat cultivars have less infertile tillers, and less early senescence than a few decades ago. They allocate more biomass to reserve and reproductive organs.
- Tiller viability and early staygreen are reaching a maximum with limited room for progress in the future.
- Further improvement could be made in transpiration efficiency, to breed for varieties which produce more crop per drop.
In Australia, wheat is mainly grown under rain-fed conditions and experiences major water stress that limits productivity (Chenu et al., 2013). Although severe water stresses are expected to slightly decrease in the Northern Region in the coming decades, drought is projected to remain a major limitation for wheat yield (Watson et al., 2015; Lobell et al., 2015).
While the physiological responses of wheat to water deficit are very complex, it is likely that traits associated with drought tolerance have been indirectly selected through conventional breeding programs in drought-prone areas. By looking at fifteen elite Australian cultivars released between 1973 and 2012, possible changes in traits related to drought adaptation were investigated. Cultivars were chosen to have with wide adoption and narrow phenological range: Suntop (2012), Scout (2010), Mace (2009), Gladius (2007), Drysdale (2002), Wyalkatchem (2001), Yitpi (1999), Krichauff (1997), Frame (1994), Janz (1989), Machete (1995), Spear (1984), Hartog (1982), Warigal (1978), Condor (1973).
Plants were grown at a density of 100 plant m-2 in the ‘Pot in Bucket’ (PIB) system adapted from Hunter et al. (2012), and had a continuous supply of water from independent water jugs, which allowed transpiration to be measured for each pot from 25 days after sowing until flowering. Phenology, plant growth and development were recorded weekly. Transpiration efficiency was calculated as the ratio between dry biomass over cumulated water transpired from 25 days after sowing until flowering.
No trending change in plant biomass, but an increased proportion of resources goes to reserve and reproductive organs.
Plant biomass did not significantly change between 1973 and 2012 (Fig. 1a) but the partitioning of that biomass changed (Fig. 1b-f). Modern varieties tended to partition more biomass to stems and spikes, at the expense of the leaves. Interestingly, the decrease in leaf biomass was a result of a decrease in dead leaves, while green-leaf biomass at flowering did not significantly change over the last decades. Hence, breeders seem to have indirectly selected for lines with reduced pre-flowering senescence that invest an increasing amount of resources towards reserve and reproductive organs, which ultimately translates in greater yield (Sadras and Lawson, 2013).
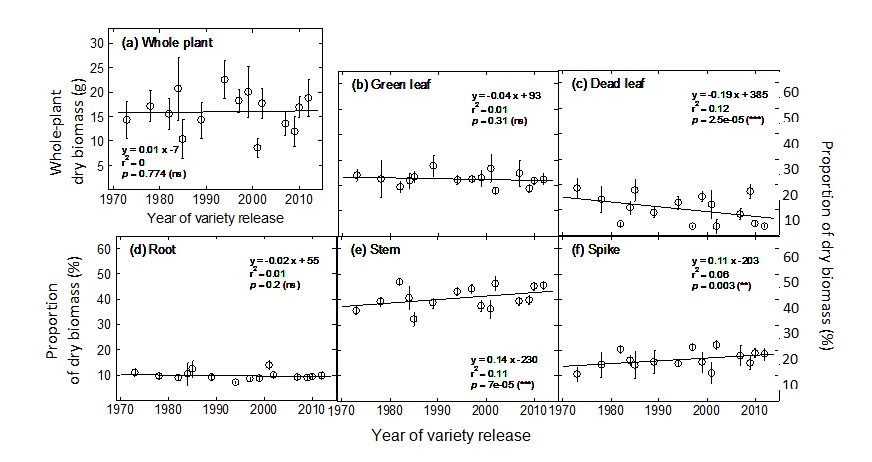
Figure 1. Change in biomass over year of variety release for (a) whole-plant dry biomass, and the portion of dry biomass allocated to (b) green leaves, (c) dead leaves, (d) roots, (e) stems, and (f) spikes at flowering. Error bars correspond to 95% confidence interval (n = 5-10). Linear regressions were calculated on row data (individual pots for root biomass or individual plants for all other traits). Abbreviations: ns, not significant; *, p < 0.05; **, p < 0.01; ***, p < 0.001.
For the varieties studied, the proportion of biomass allocated to the stems and the spikes at flowering has increased by +0.14% and +0.11% per year, respectively. The most noticeable effect was for the spikes which increased from 13 to 18% of total plant biomass over the last four decades (Fig. 1f). For comparison, the stem portion increased from 37% to 42% during the same period (Fig. 2e).
This change in partitioning can be at least be partly explained by a change in tillering (Fig. 2). While the number of total tillers has slightly but significantly decreased, the number of fertile tillers has substantially increased (from 5.5 to 7.0 between 1970 and 2010; Fig. 2b). Accordingly, the number of infertile tillers has drastically and very significantly decreased to the point that most modern varieties have almost no infertile tillers (Fig. 2c).
Overall, the changes observed in tillering, senescence and biomass partitioning reflect that modern varieties (i) invest less resources in plant structure that die relatively early during the crop cycle, but (ii) accumulate more biomass in reproductive organs (spike) and in reserve tissues (stems), which can benefit yield under late drought conditions (Dreccer et al., 2009).
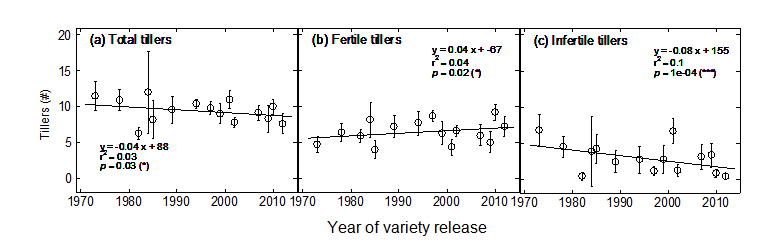
Figure 2. Change in (a) tiller number, (b) fertile tillers, and (c) infertile tillers at flowering, over year of release. Error bars correspond to 95% confidence interval (n = 10). Linear regressions were calculated on row data (individual plants). Abbreviations: ns, not significant; *, p < 0.05; **, p < 0.01; ***, p < 0.001.
Transpiration efficiency has significantly increased between 1973 and 2012.
While modern cultivars seem more efficient in partitioning their resources towards reproductive organs, they were also found to be more efficient in converting transpired water into plant biomass (Fig. 3). Plant transpiration efficiency (Fig. 3) has increased significantly with the year of release.
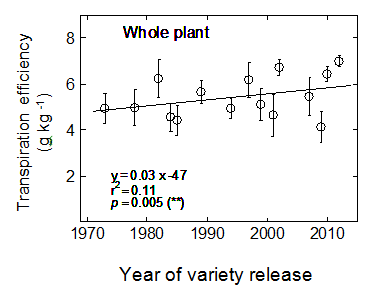
Figure 3.Change in transpiration efficiency for whole-plant dry biomass for varieties released between 1973 and 2012. Error bars correspond to 95% confidence interval (n = 10). Linear regressions were calculated on row data (individual plants). Abbreviations: ns, not significant; *, p < 0.05; **, p < 0.01; ***, p < 0.001.While modern cultivars seem more efficient in partitioning their resources towards reproductive organs, they were also found to be more efficient in converting transpired water into plant biomass (Fig. 3). Plant transpiration efficiency (Fig. 3) has increased significantly with the year of release.
Conclusion
While this study was conducted under well-watered conditions, it revealed substantial changes in traits likely to be beneficial under drought conditions. A significant trend in increasing number of fertile tillers was observed with the year of release, while the number of total tillers and infertile tillers significantly decreased. Modern cultivars had less early senescing leaves but maintained a similar plant leaf area and green-leaf biomass to older varieties at flowering. Most importantly, a significant increase in transpiration efficiency was observed for modern varieties. To put these results into perspective, during the 70s wheat varieties were producing approximately 4.8g of biomass for every litre of water transpired, while modern varieties have increased biomass accumulation to just over 5.8g per litre. Overall, modern wheat cultivars were able to use approximately 500g less water per plant up to flowering, which for a density of 100 plants m-2 corresponds to a saving of 50 mm of water. In the Northern Region, such pre-flowering water saving would result in the field in a greater reserve of soil water for use during grain filling, thus allowing to reduce terminal drought.
Promising variability exists for transpiration efficiency and could be used to assist breeders developing lines that produce more crop per drop.
Acknowledgements
This study was funded by the University of Queensland and the ARC Centre of Excellence for Translational Photosynthesis, with some support from GRDC projects.
References
Chenu, K., Deihimfard, R. & Chapman, S. C. (2013). Large-scale characterization of drought pattern: a continent-wide modelling approach applied to the Australian wheatbelt spatial and temporal trends. New Phytologist 198(3): 801-820.
Dreccer, M. F., van Herwaarden, A. F. & Chapman, S. C. (2009). Grain number and grain weight in wheat lines contrasting for stem water soluble carbohydrate concentration. Field Crops Research 112(1): 43-54.
Hunter, M., Mitchell, J. & Dieters, M. (2012).Semi-automated, non-weighing, pot-in-bucket (PIB), water management in pot plant culture. In 16th Australian Agronomy ConferenceArmidale, NSW, Australia: Australian Society of Agronomy.
Lobell, D. B., Hammer, G. L., Chenu, K., Zheng, B., McLean, G. & Chapman, S. C. (2015). The shifting influence of drought and heat stress for crops in northeast Australia. Glob Chang Biol 21(11): 4115-4127.
Sadras, V. O. & Lawson, C. (2013). Nitrogen and water-use efficiency of Australian wheat varieties released between 1958 and 2007. European Journal of Agronomy 46(0): 34-41.
Watson, J., Zheng, B., Chapman, S. & Chenu, K. (2015).Impact of projected climates on drought occurrence in the Australian wheatbelt. In 17th Australian Agronomy Conference, 4 Hobart, Australian.
Contact details
Dr Karine Chenu
The University of Queensland, Queensland Alliance for Agriculture and Food Innovation (QAAFI)
203 Tor Street
PO Box 102
Toowoomba, QLD 4350
Ph: 07 4688 1357
Fx: 07 4688 1193
Email: karine.chenu@uq.edu.au
Varieties displaying this symbol beside them are protected under the Plant Breeders Rights Act 1994